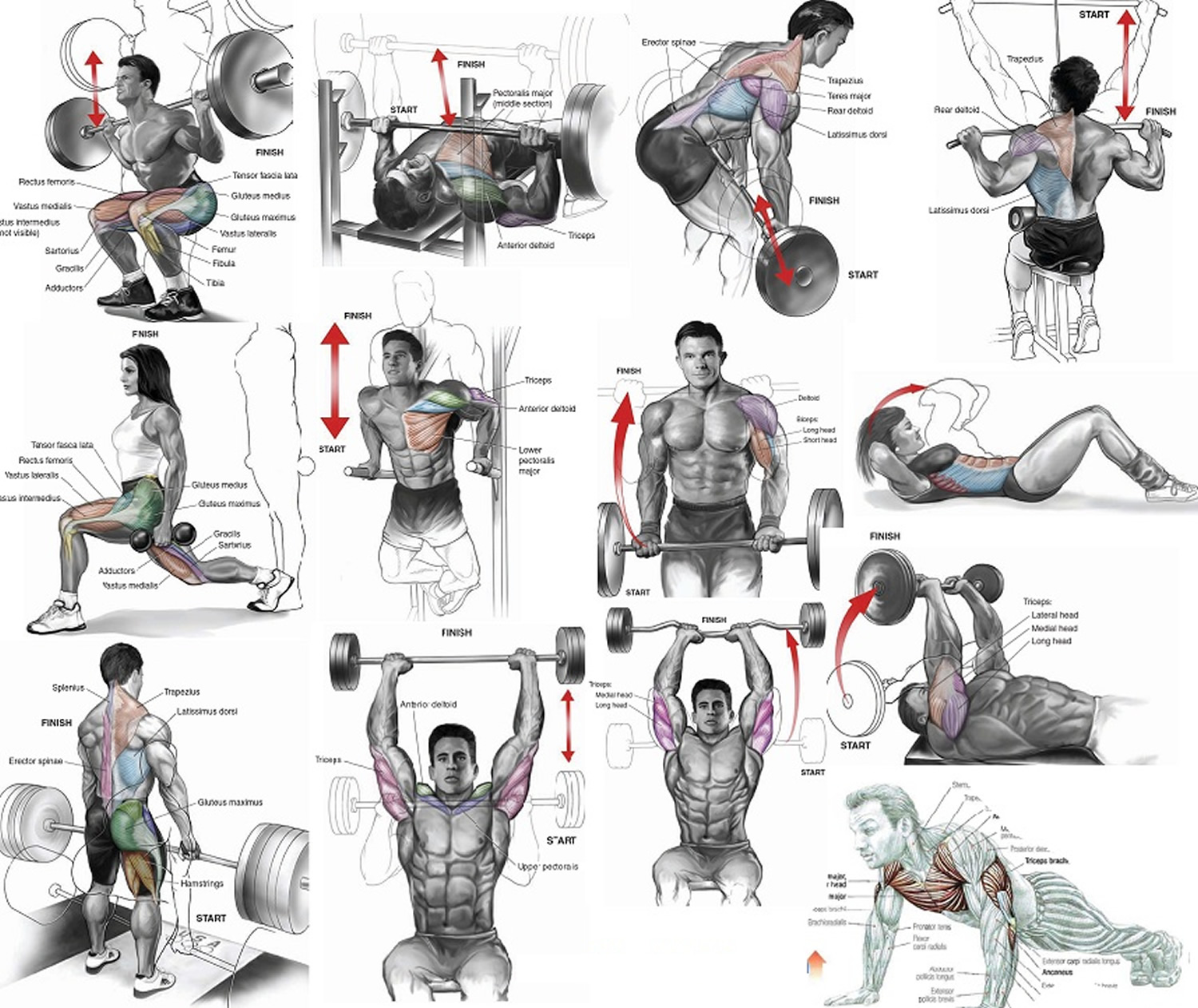
What are muscles ?
Did you know you have more than 600 muscles in your body ? These muscles help you move, lift things, pump blood through your body, and even help you breathe.
When you think about your muscles, you probably think most about the ones you can control. These are your voluntary muscles, which means you can control their movements. They are also called skeletal muscles, because they attach to your bones and work together with your bones to help you walk, run, pick up things, play an instrument, throw a baseball, kick a soccer ball, push a lawnmower, or ride a bicycle. The muscles of your mouth and throat even help you talk and eat.
Smooth muscles are also called involuntary muscles since you have no control over them. Smooth muscles work in your digestive system to move food along and push waste out of your body. They also help keep your eyes focused without your having to think about it.
Did you know your heart is also a muscle ? Cardiac muscle is a specialized type of involuntary muscle. It pumps blood through your body, changing its speed to keep up with the demands you put on it. It pumps more slowly when you’re sitting or lying down, and faster when you’re running or playing sports and your skeletal muscles need more blood to help them do their work.
Skeletal muscles are connected to your bones by tough cords of tissue called tendons. As the muscle contracts, it pulls on the tendon, which moves the bone. Bones are connected to other bones by ligaments, which are like tendons and help hold your skeleton together.
Skeletal muscle is a highly plastic tissue. The ability of adult muscle fibres to change in response to external stimuli has been called muscle plasticity. Force, contraction speed, endurance and oxidative/glycolytic capacity are all examples of muscle properties that are plastic 1. Skeletal muscle is a permanent, post-mitotic tissue, and unless the muscle is damaged there is little turnover of cells 2, 3. Thus, it has been demonstrated that dramatic changes in gene expression, protein composition and physiological properties can occur in pre-existing fibres without de- or regeneration 4, 5. The plastic changes occur mainly by reprogramming the cell by turning on and off sets of relevant genes.
Exercise evokes signaling pathways that strongly modify myofiber metabolism and physiological and contractile properties of skeletal muscle. Regular physical activity is beneficial for health and is highly recommended for the prevention of several chronic conditions 6.
Muscle fibre phenotypes
Mammalian skeletal muscles are composed of myofibers with various contractile properties (such as force production, endurance, twitch duration, and shortening velocity) and differing metabolism. Myofibers are mainly classified as slow-twitch or fast-twitch based on the maximal speed of shortening 7. In human muscle 3 fibre subtypes are recognized based on their contractile and metabolic properties 8. Fast-twitch fatigable fibres rely predominantly on glycolytic metabolism and are designated FG (fast glycolytic), whereas, fast-twitch fatigue-resistant and slow-twitch fibres have relatively greater mitochondrial content and are designated FOG (fast oxidative glycolytic) and SO (slow oxidative), respectively 9. The fibre type-specific differences in contractile function are due to differential expression of a diverse range of isoforms of each myofibrillar protein 10. Myosin heavy chain (MyHC) isoforms are intimately associated with myofibre contractile and energetic properties and are commonly used molecular markers: FG fibres express MyHC IIx/d, FOG fibres express MyHC IIa and SO fibres express MyHC I, which is also the predominant isoform in adult human myocardium. Indeed, myosin ATPase activity determines the sliding velocity between actin and myosin, thereby shortening the velocity of the fiber 11. Myosin ATPase type I histochemical staining identifies slow-twitch fibers, while myosin ATPase type II (which has the highest ATPase activity) stains fast-twitch myofibers. Based on the expression of the predominant isoforms of MyHC protein expressed, myofibers are mainly classified as type I fibers, type IIx/d fibers, and type IIa fibers 7, 12 (Figure 1).
In addition to the three or four major MyHC genes expressed in adult limb muscles there are specialized forms expressed during development and in gill-arch-derived muscles. In total 10 different MyHC genes have been connected to skeletal muscle 13.
While histochemical or immunohistochemical staining might give the impression that the vast majority of fibres are positive only for one MyHC, single-fibre gel electrophoresis has revealed that 11–67% of the fibres from various limb muscles express more than one MyHC isoform even under steady-state activity conditions 14. It can be concluded that the concept of universal fibre types throughout the body is an oversimplification.
Figure 1. Characteristics of mammalian skeletal muscle fiber types. The red color is associated with a high content of myoglobin
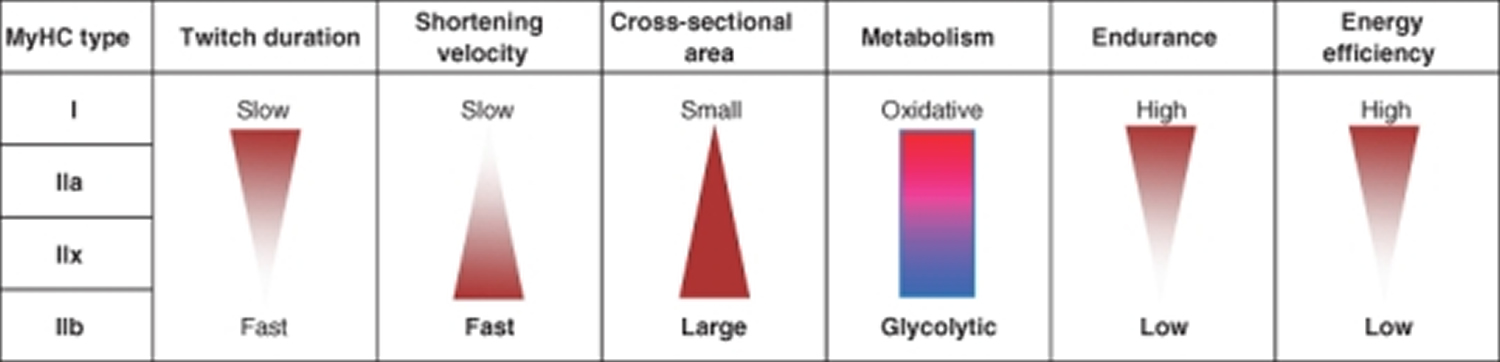
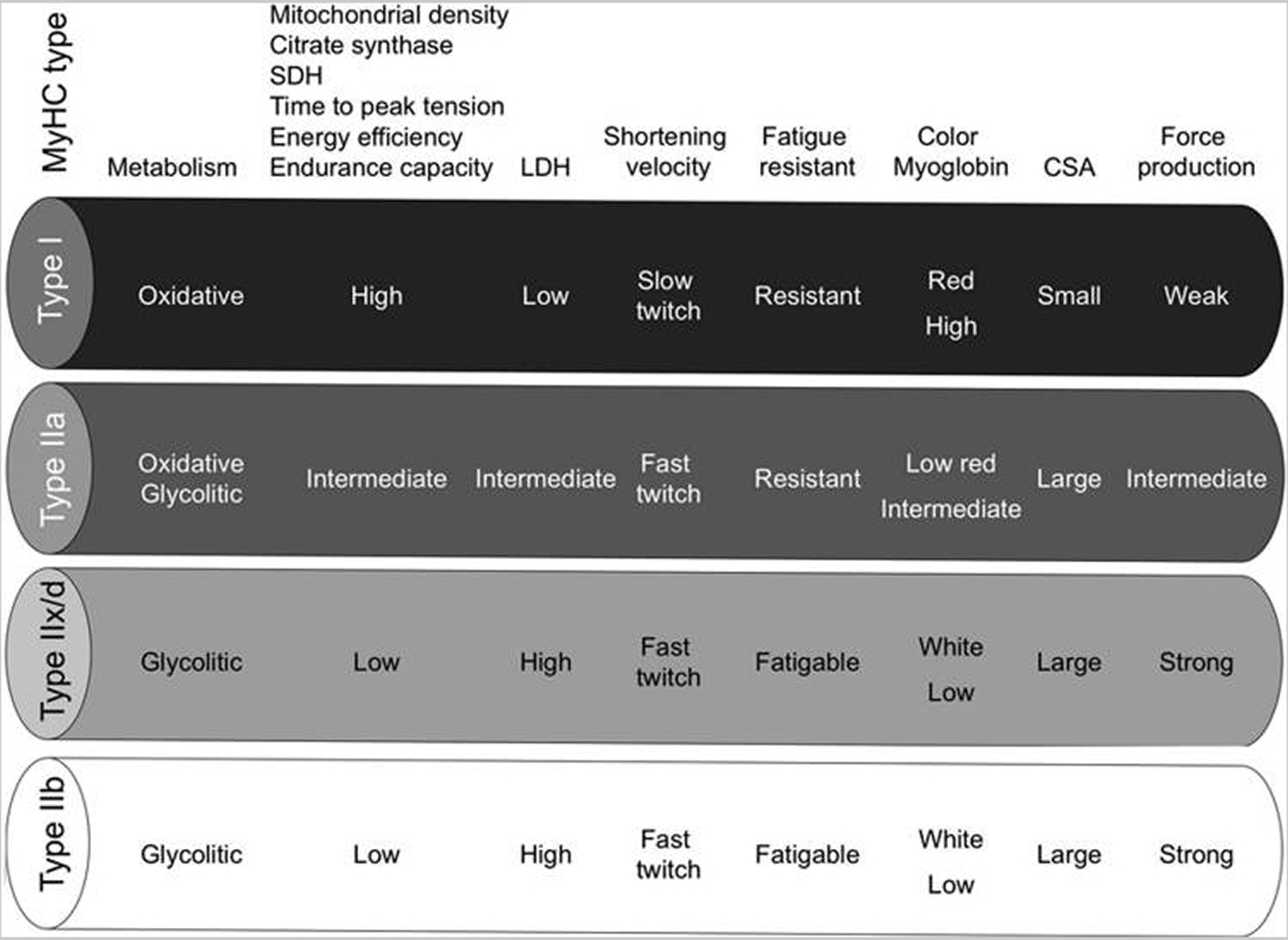
Note: MyHC, myosin heavy-chain; SDH, succinate dehydrogenase; LDH, lactate dehydrogenase; CSA, cross-sectional area.
Type I fibers (slow-twitch fibers) contain the slow isoform of MyHC and slow isoforms of other contractile proteins. They have a predominatly oxidative metabolism. They are characterized by high mitochondrial content, high capillary density and express mainly glucose and fatty acid oxidative enzymes. Type I fibers are rich in myoglobin and are red colored. They develop a slow contractile force and are resistant to fatigue. They are involved in continuous tonic activity. Force production depends on the time the myosin head spends bound to actin, on the myosin head density and on the duty ratio 15.
Type IIx/d fibers (fast-twitch fibers) express a fast isoform of MyHC and fast isoforms of other contractile proteins and, therefore, develop a fast contractile force. Type IIx/d fibers mainly metabolize glucose by glycolysis and are characterized by low mitochondrial content and low capillary density. They are also poor in myoglobin and are white in appearence. Type IIx/d fibers express low glucose transporter 4 (GLUT4) and have low sensitivity to insulin that type I fibers. They are involved in phasic activity 16.
Type IIa fibers (fast-twitch fibers) have intermediate features. They have a mixed (oxidative/glycolytic) metabolism. They are fast-twitch fibers with a fast contractile force development, but mainly express oxidative enzymes. Although muscle endurance and resistance to fatigue rely on several cellular factors, there is a strict correlation between these properties and high oxidative capacity and high content of mitochondria of the fiber. Therefore, type IIa fibers are fast but they are more resistant to fatigue than type IIx/d fibers as they are more oxidative 17, 18.
Rodents also possess type IIb fibers that are more fast-twich and glycolytic than IIx/d fibers (Fig. 1). Many other contractile and structural proteins are also present in distinct isoforms whose expression is more or less tightly connected to fiber type. For example, the shortening velocity also depends on myosin light chain isoforms; thus, it might vary among fibers of the same MyHC type. Therefore, the classification reported earlier in four main fiber types is an oversimplification. Moreover, muscle also contains hybrid fibers with a combination of myosin transcripts (I-IIa-IIx/d-IIb).
The velocity of shortening and the fiber’s twitch duration depend not only on myosin composition but also on the speed of Ca2+ release and uptake in the fiber. These, in turn, depend on the development of sarcoplasmic reticulum and on sequestering systems such as the sarcoplasmic reticulum Ca2+ ATPases (SERCAs) whose isoforms are differentially expressed in different fiber types 17, 19.
Changes in muscle fibre phenotypes
The physiological properties (shortening velocity, twitch duration, endurance, etc.) that are linked in a fibre type are related to highly different molecular families (MyHC, SERCA, metabolic enzymes, etc.). Coupling regulation of different physiological properties may be beneficial from an energy conservation point of view, and/or it might reflect common signaling systems diverging to regulate several sets of diverse genes. To some extent however, different properties can be uncoupled and regulated independently during plastic changes. For example, some degree of uncoupling has been observed between twitch speed and shortening velocity 20. More importantly, endurance-exercise in man and in other animals can lead to pronounced changes in metabolic properties without MyHC fibre-type conversion 21, 22, 23, although exercise can also change MyHC type in particular within type II (e.g. from type IIb/IIx to IIa) 24, 25, 26 and under more severe experimental conditions fibre-type conversions are frequent.
When fibre-type conversions occur, it usually happens in a sequential order 27, 10: I ↔ IIa ↔ IIx ↔ IIb. During transitions hybrids between the “nearest neighbour” fibre type in this flow chart (e.g. I+IIa, IIa+IIx) are common, but aberrant hybrids such as I+IIb, I+IIa+IIb and I+IIx+IIb can also be seen under some experimental conditions 28.
Determinants of muscle fibre phenotype
The factors that determine the molecular make-up of already formed adult muscles, and how that make-up can change. At any point in time, a fibre’s make-up appears to depend on previous: (1) cell history/lineage; (2) nerve-evoked electrical activity; (3) mechanical conditions; (4) para-/autocrine conditions; and (5) circulating hormones.
There is a consensus that changes in muscle usage will transform muscle phenotype, but the precise biological signaling mechanisms responsible for such changes are less clear.
Muscle properties are strongly influenced by hormones such as testosterone and thyroid hormones, as reviewed previously 29, 18, 30. The link between external factors related to activity and usage (points 2 and 3 above) and gene expression and the ability to change is, however, constrained by the muscle’s cell lineage.
The importance of cell lineage
Developmental studies suggest that initial fibre-type differentiation might be determined by myoblast cell lineage independent of external influences such as innervation or usage.
In adult rats, when different muscles are regenerating from myoblasts after myofibre destruction, the various regenerated muscles express different MyHC types reminiscent of the muscle of origin. This happens even if the muscles receive similar experimental patterns of activity. Thus, when regenerating soleus and extensor digitorum longus (EDL) were stimulated by the same slow pattern, the EDL failed to express the large amount of slow MyHC that was observed in the soleus under the same conditions 31.
While it seems clear that cell lineage limits the adaptive range of muscle plasticity, it is equally clear that external signals can change muscle phenotype in the adult. In particular signals from the nerve appear to be important.
It can be concluded that muscle fibre pedigree matters, and that there is a cell line component to the resulting adult phenotype of a fibre. The relationship is, however, not simple, since experiments with genetically marked myoblasts suggest that single myoblast clones can contribute to both fast and slow fibres, clones are not restricted to contribute to subsets of fibre types, and clones show no detectable preference for fusion to a particular fibre type 32.
What are the signals from the nerve ?
There is currently no compelling evidence to suggest that there are any relevant sources of neural influence on the muscle other than activity, and in spite of intense searching for several decades, no neurotrophic substances have been identified that prevent atrophy or mimic other effects of normal innervation or cross-innervation outside the synaptic zone 1.
The importance of nerve-evoked muscle activity
Generally type I motor units seem to receive high amounts of impulses delivered in long low-frequency trains, while type II units seem to receive short bursts of high-frequency activity. The total amount of impulses delivered to type II units is lower, but the amount seems to vary among the type II subtypes.
Mechanical stress
It is widely assumed that contraction against a resistance leads to larger muscles than contraction against lower resistance, but this does not necessarily have a direct bearing on the importance of mechanical factors as such.
The most compelling evidence for a mechano-dependent mechanism comes from experiments where limbs have been immobilized by a cast. This leads to atrophy, but studies over almost 100 years have shown that atrophy can be partly counteracted when muscles are immobilized in a lengthened position rather than a shortened position. There are also studies suggesting that muscle length influences contraction speed such that chronic stretch makes a muscle slower; immobilization of fast muscles in a lengthened position thus increases the fraction of slow fibres. In most experimental conditions it is hard to separate electrical activity and mechanical stretch, but some experimental data point to the presence of an activity-independent mechanical mechanism influencing muscle size and perhaps contraction speed.
Myostatin
Myostatin (previously called GDF-8) is a member of the transforming growth factor β (TGF-β) superfamily and plays a major role during development where it acts as an inhibitor of muscle growth. Disruption of the myostatin gene leads to development of grossly enlarged muscles in mice 33, farm animals 34, and man 35. The enlargement is caused both by an increase in the number of fibres (hyperplasia) and in fibre size (hypertrophy). Importantly, muscle enlargement obtained by myostatin deficiency is peculiar because it does not increase force in proportion to size, thus the amount of contractile proteins may not be properly regulated 36, 37. Thus, reducing myostatin alone might not mimic effects of strength training, although strength training in adults has been shown to be associated with reduced levels of myostatin in muscle and plasma 38, 39. In adult animals inhibition of myostatin with antibodies leads to hypertrophy without an increase in the number of fibres 40; conversely, overexpression of myostatin in muscle fibres after electroporation leads to muscle atrophy without loss of muscle fibres 41. In the latter study it was suggested that myostatin acted by reducing muscle gene expression of myofibrillar proteins perhaps by reducing expression of MyoD and myogenin. In addition, myostatin might activate the ubiquitin-proteasome pathway for proteolysis 42.
Insulin-like growth factor I (IGF-1)
IGF-1 has been implicated as a factor promoting hypertrophy in the adult animal. The liver supplies approximately 75% of the circulating IGF-1 (Schwander et al., 1983), and a selective abolishment of IGF-1 production in hepatocytes leads to a 75% reduction in circulating IGF-1 levels but without growth impairment 43. In humans increasing the circulating level of IGF-1 does not promote muscle protein synthesis 44. IGF-1 is also expressed locally in several tissues including muscle where it is induced by stretch or high-resistance exercise. IGF-1 seems to work as a local hormone that promotes hypertrophy in adult muscle. It might do so both by interfering with protein balance in muscle fibres and by activating satellite cells, but for the latter there is still little information in adult muscles.
Figure 2. Muscles anatomy front
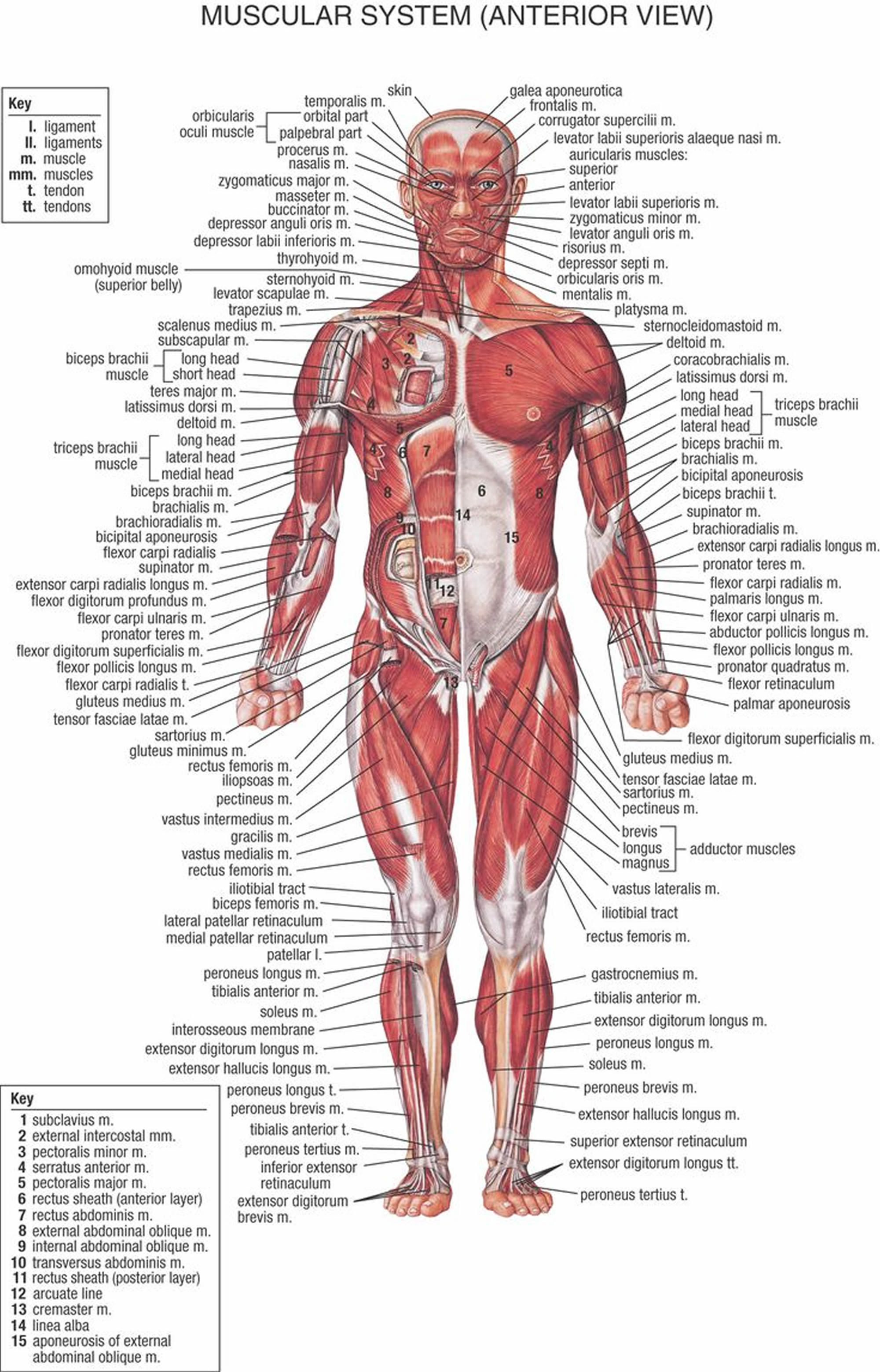
Figure 3. Muscles anatomy back
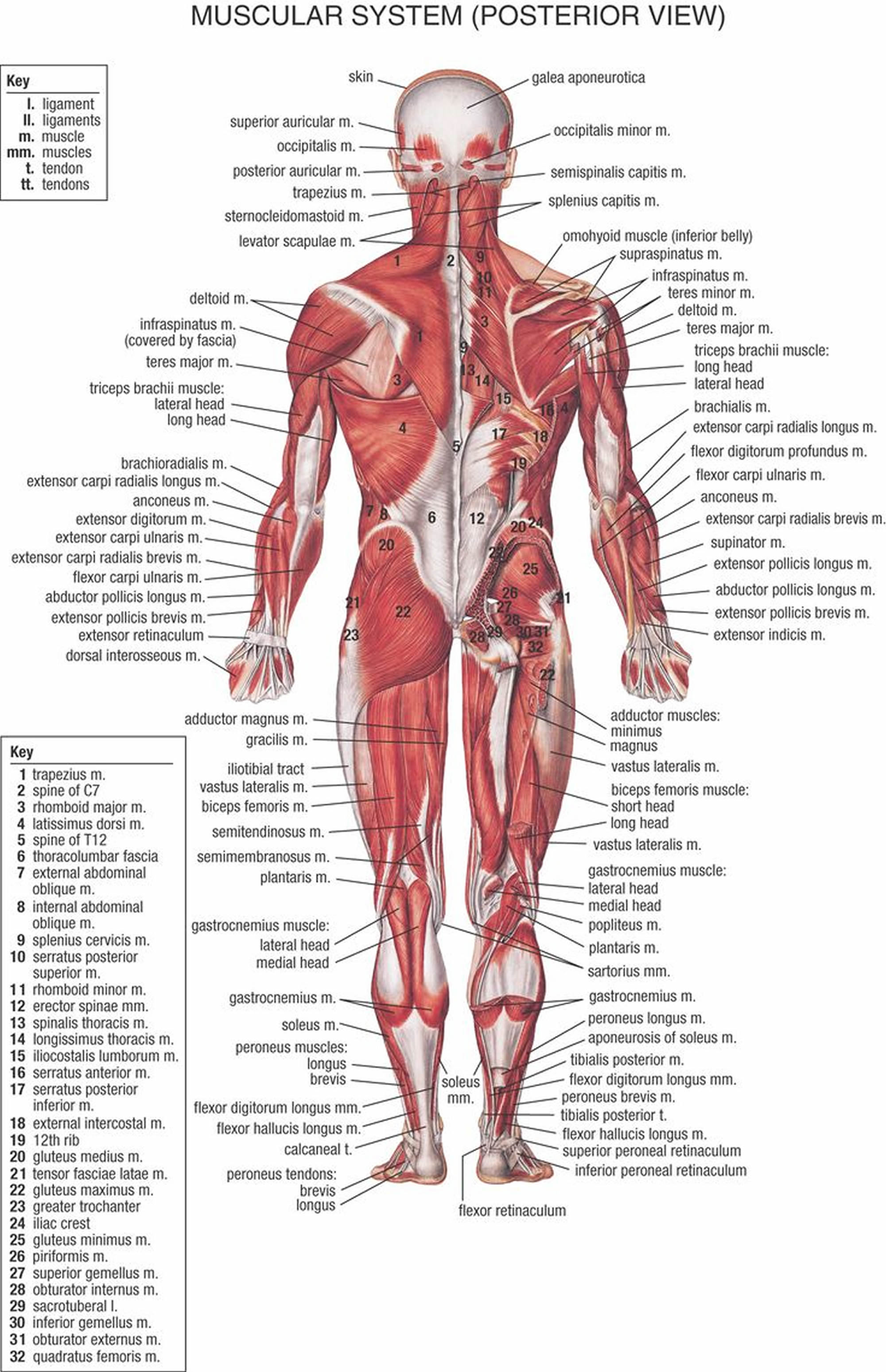
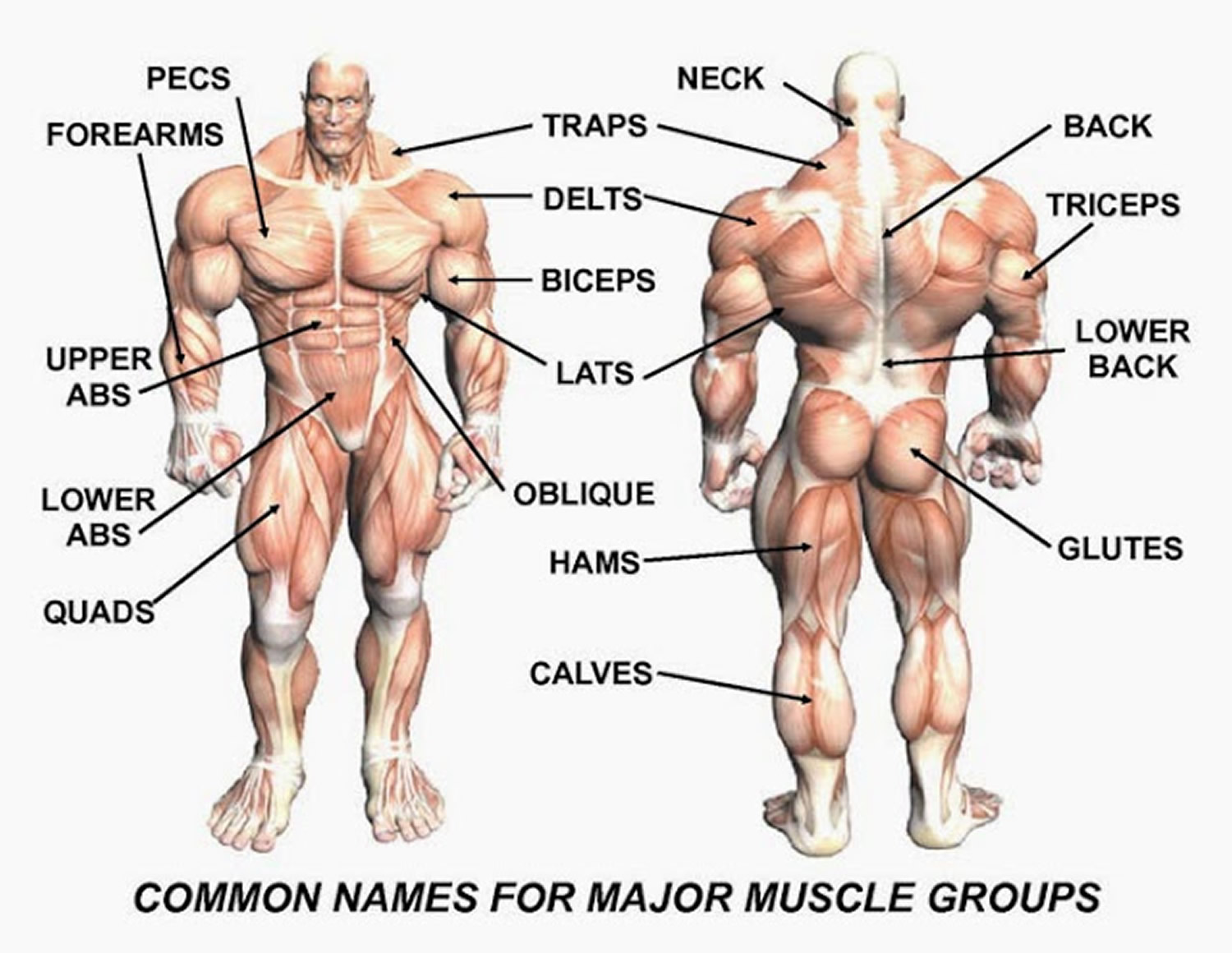
Figure 4. Muscle anatomy – simplified for bodybuilding and body builders
The cell biology of muscle fibre size
Regulation of force is mainly a question of regulating fibre size, and ultimately size is regulated by altering the balance between protein synthesis and degradation in each muscle fibre. Change in fibre size can be achieved by regulating three major conditions: (1) the number of nuclei; (2) the rate of protein synthesis for each nucleus; and (3) the rate of protein degradation.
Figure 5. Pathways currently believed to be involved in regulating muscle fibre size
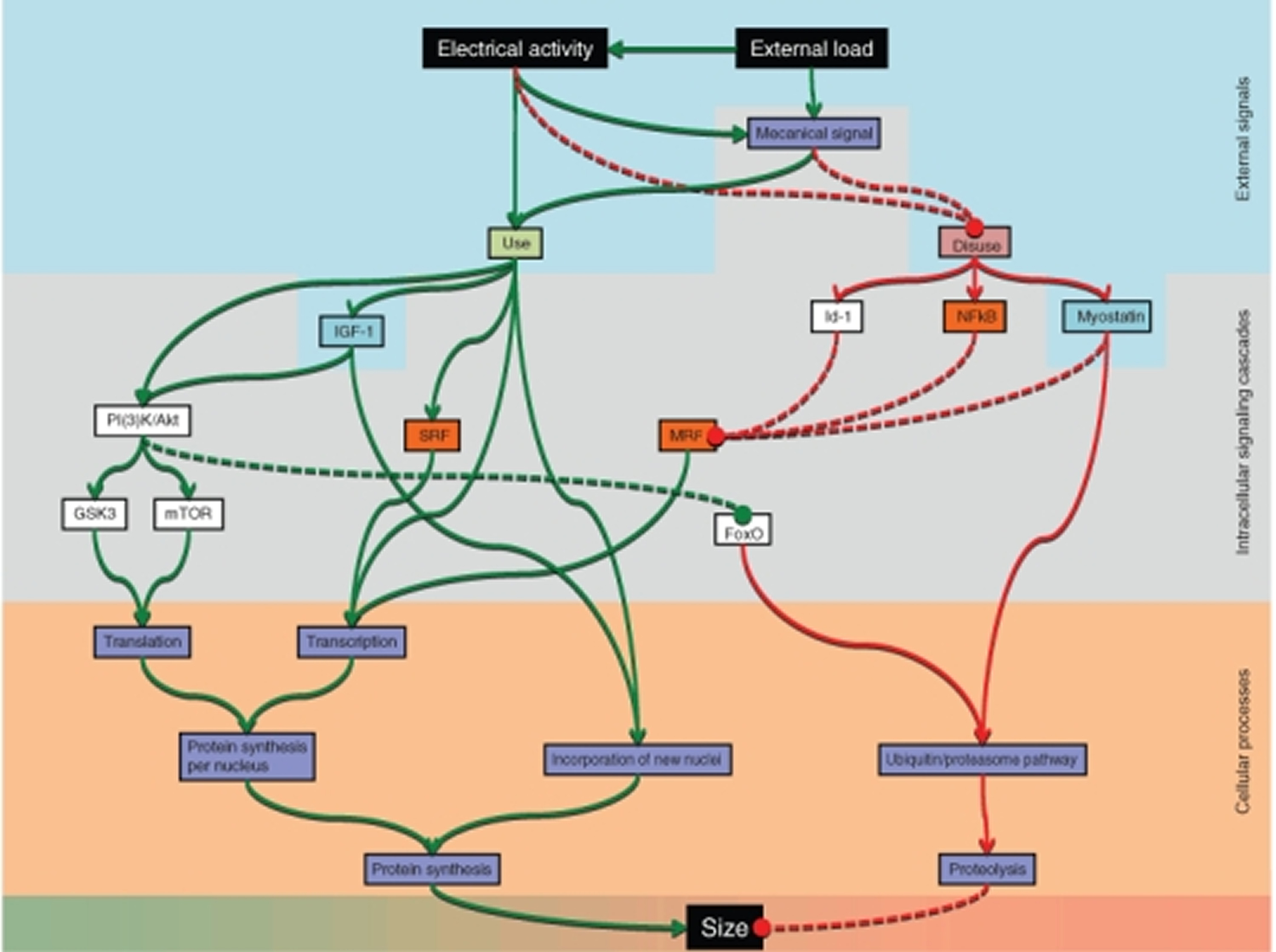
The pathways have different degrees of scientific support, and their relative importance is still poorly understood. Abbreviations: forkhead box O (FoxO), glycogen synthase kinase 3 (GSK3), inhibition-of-DNA-binding-protein 1 (Id-1), insulin-like growth factor I (IGF-1), mammalian target of rapamycin (mTOR), myogenic regulatory factor (MRF), phosphatidylinositol 3-kinases/Akt (PI(3)K/Akt), serum response factor (SRF), κ light polypeptide gene enhancer in B-cells (NFκB).
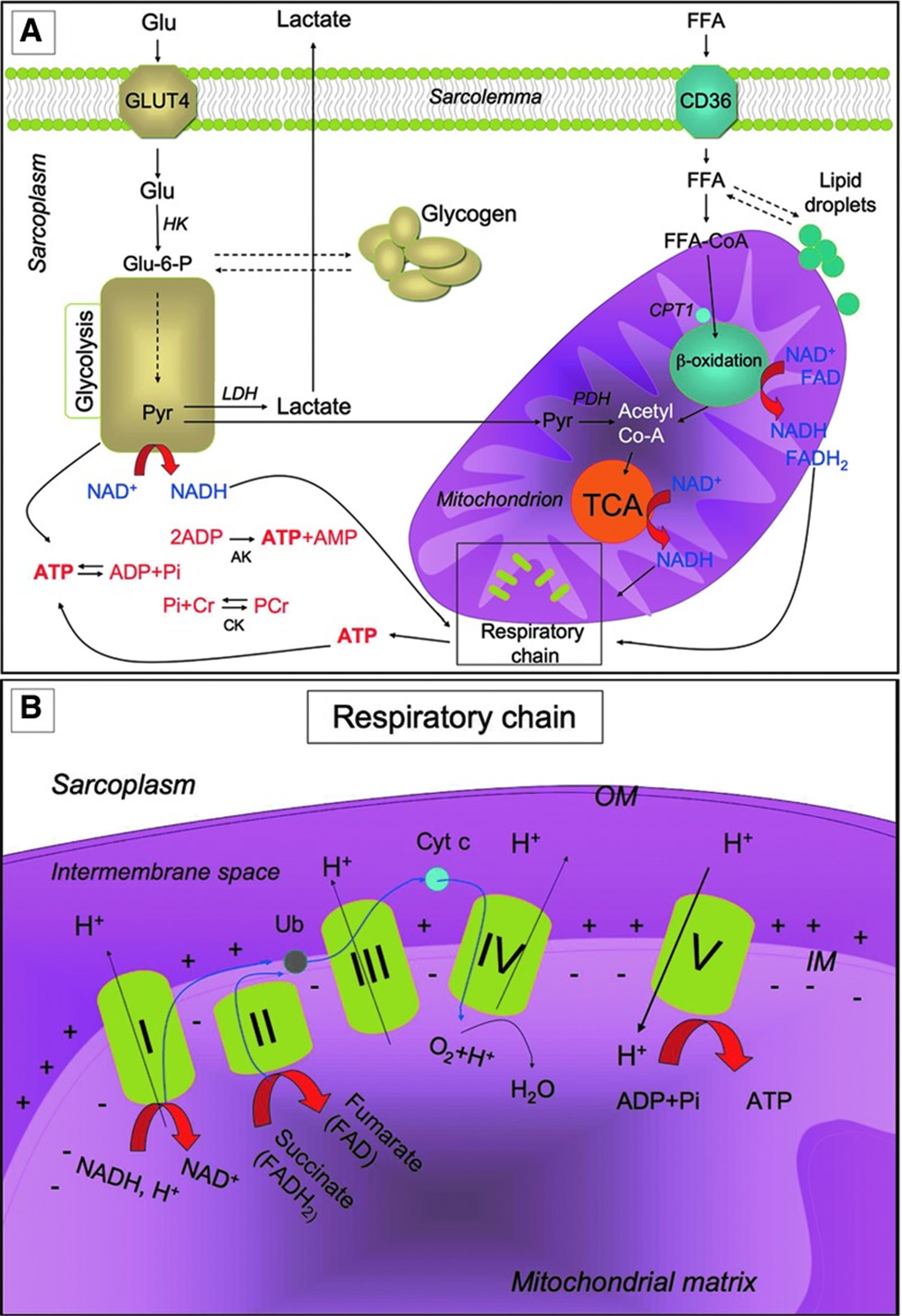
Metabolic pathways for ATP production in skeletal myofibers
Metabolic pathways for ATP production in skeletal myofibers. (A) Skeletal muscles require a high amount of ATP for contraction. The main sources of energy are Glu and FFA. Glu uptake into the sarcoplasm from blood occurs, among other things, through the GLUT4. Once in the cytosol, Glu is phosphorylated by HK and forms Glu-6-P. One molecule of Glu-6-P can be converted into two molecules of Pyr through glycolysis, a metabolic anaerobic pathway involving 10 enzymes (the enzyme phosphofructokinase is an important control point in the glycolytic pathway). Depending on the energy needs, Glu-6-P can also be stored as glycogen. In anaerobic conditions Pyr is reduced to lactate by LDH. Alternatively, in aerobic conditions, Pyr might be transferred into the mitochondria matrix, where it is decarboxylated into acetyl-CoA by the PDH complex. Acetyl-CoA is then metabolized through the TCA cycle. The first enzyme acting in the TCA cycle is the citrate synthase that forms citrate from acetyl-CoA and oxaloacetate. The TCA cycle produces reducing equivalents (NADH, FADH2) and CO2. In addition to Pyr, another important source of acetyl-CoA is the β-oxidation of FFA. FFA enter the myofiber through a passive flip-flop or through a protein-mediated mechanism such as the FAT/CD36. In the cytosol, FFA undergo esterification and form triglycerides stored as lipid droplets that are surrounded by mitochondria. Alternatively, at the mitochondrial OM, they can be condensed with CoA to form FFA-CoA and, through the CPT1, they can cross the mitochondrial IM and reach the mitochondrial matrix where they undergo β-oxidation. β-oxidation is a cycle of four reactions. Each cycle produces a molecule of acetyl-CoA which, in turn, enters the TCA cycle. Along with acetyl-CoA, during β-oxidation, FADH2 and NADH are also formed. In skeletal muscles, at rest, excess of ATP produced is stored as PCr. ATP is converted into ADP and Pi by ATPase, and the Pi is used to convert Cr in PCr whose amount is roughly 10 times higher than the amount of ATP. During intense activity, PCr can anaerobically donate a phosphate group to ADP and form ATP for quick regeneration of ATP. PCr is, therefore, a rapid system to supply energy during contraction. The reversible phosphorylation of Cr is catalyzed by several CK. Once ATP also produced by PCr is consumed, the AK (myokinase) catalyzes the formation of ATP and AMP from two ADP molecules. During exercise, the amount of ATP produced by the myofiber increases enormously. However, the stores of ATP that can be detected in the myofiber are not as high, as ATP is stored in the form of PCr. (B) Reducing equivalents (NADH and FADH2) generated mainly during TCA, β-oxidation, and glycolysis are oxidized by the complexes of the respiratory chain (Complex I, II, III, and IV) in the oxidative phosphorylation pathway. Electrons are transferred from NADH and FADH2 to oxygen (which is reduced to H2O) by means of the enzyme complexes and by the electron carriers Ub and Cyt c of the respiratory chain. The energy released by reducing equivalent oxidation as electrons pass from one complex to the next is used to pump protons (H+) across the IM into the intermembrane space. This creates an electrochemical proton gradient across the IM, which is highly energetic. Protons can flow along this gradient through ATP synthase (ATPase or complex V); this backflow releases the energy of the proton gradient, which is used by ATP synthase to phosphorylate ADP and to form ATP. This phosphorylation of ADP is called oxidative, as it is coupled to the presence of oxygen that enables the oxidation of reducing equivalents. By this mechanism, nutrients are oxidated and their energy is stored in usable energy as ATP. ATP is also produced in a lower amount during glycolysis. OM, outer membrane; Glu, glucose; FFAs, free fatty acids; Glu-6-P, glucose-6-phosphate; Pyr, pyruvate; PDH, pyruvate dehydrogenase; TCA, tricarboxylic acid; FAT/CD36, fatty acyl translocase; CPT, carnitine palmitoyltransferase; CK, creatine kinases; AK, adenylate kinase; Ub, ubiquinon; IM, inner membrane; Cr, creatine; Cyt c, cytochrome c; GLUT4, glucose transporter 4; HK, hexokinase; PCr, phosphocreatine.
Figure 6. Muscle metabolism
Exercise-Induced Adaptation
Skeletal muscle is extremely adaptable to environmental changes and is characterized by a high metabolic flexibility: It is able to rapidly modify the rate of ATP synthesis, the blood flow, and the kind of substrate used, depending on needs 45. Skeletal muscle is also extremely adaptable to changes in contractile activity: Physical exercise strongly modifies metabolic potential, morphology, and physiology of skeletal muscle, thus producing a strong beneficial effect on health 46. All pathways of ATP generation are active during exercise, but the relative contribution of each is determined by the intensity and duration of contraction. Indeed, exercise might be performed with different modalities, thus producing different effects on muscles 47.
Physical exercise might be grossly classified as “endurance training” and “resistance training.” Endurance training is based on endurance and is aerobic, while resistance training is based on strength. Endurance exercise (e.g., performed by marathon runners, swimmers, and cyclists) is generally characterized by high-frequency, long duration, and low power output. Resistance exercise (e.g., body building and throwing events) is, in general, characterized by low frequency, high resistance, high intensity, and short duration. Along with the modality of exercise, other parameters such as duration, frequency, and intensity of the exercise influence the effect of physical training on the muscle 48.
Exercise triggers a metabolic and structural remodeling in skeletal muscle, thus leading to changes in contractile properties and to increased angiogenesis in order to reduce muscle fatigue. These adaptations improve skeletal muscle performance. The specific features of skeletal muscle adaptation to exercise depend on the modality of exercise performed. Resistance exercise acts mainly by increasing muscle mass and strength (see “Exercise and skeletal muscle mass” section). On the other hand, endurance exercise stimulates mitochondrial biogenesis and expression of mitochondrial respiration and FFA β-oxidation genes, thereby providing a phenotypic adaptation toward a more oxidative phenotype. Submaximal aerobic activities increase insulin-independent glucose uptake and utilization in skeletal muscle, along with insulin sensitivity and redistribution of GLUT4 to the plasma membrane 49. With regard to the contractile properties, endurance exercise promotes fiber type transformation toward the slow-twitch contractile apparatus by inducing a dramatic modification of gene expression and physiological properties of the myofiber. The muscle used frequently needs to be more energy efficient, with both longer twitches and slower MyHC types contributing to higher energy efficiency 50.
Exercise provides numerous beneficial effects on skeletal muscle and, in general, on health. Although both exercise modalities are beneficial for health, endurance exercise is more effective for preventing cardiovascular diseases; while resistance training (mostly inducing muscle hypertrophy) is more effective for the maintenance of muscle mass contrasting atrophy and age-related muscle wasting 51.
Exercise and skeletal muscle mass
As stated earlier, while endurance exercise acts by up-regulating mitochondrial metabolism and fiber-type transformation, the beneficial effects of resistance exercise mainly depend on its ability to increase muscle mass. Muscle fiber hypertrophy was a determinant of overall muscle enlargement as a result of resistance training. Although many training variables contribute to the performance, cellular and molecular adaptations to resistance exercise, relative intensity (% 1 repetition maximum [%1RM]) appears to be an important factor 52. This review summarises and analyses data from numerous resistance exercise training studies that have monitored percentage fibre type, fibre type cross-sectional areas, percentage cross-sectional areas, and myosin heavy chain (MyHC) isoform expression. In general, relative intensity appears to account for 18-35% of the variance for the muscle hypertrophy response to resistance exercise. On the other hand, fibre type and MyHC transitions were not related to the relative intensity used for training. When competitive lifters were compared, those typically utilising the heaviest loads (> or =90% 1RM), that is weightlifters and powerlifters, exhibited a preferential hypertrophy of type II fibres when compared with body builders who appear to equally hypertrophy both type I and type II fibres. These data suggest that maximal hypertrophy occurs with loads from 80-95% 1RM 52.
Skeletal muscle mass depends on a delicate balance between protein synthesis and protein degradation: Resistance exercise influences both these processes by activating the phosphoinositide 3-kinase (PI3K)/Akt/mammalian target of rapamycin (mTOR) signaling 53. The kinase mTOR exists in two independent complexes: mTOR complex 1 (mTORC1) and mTORC2. Raptor and Rictor are specific functional components of TORC1 and TORC2, respectively. mTORC1 controls protein translation by phosphorylating the eukaryotic translation initiation factor 4E-binding protein-1 (4E-BP1) and p70 ribosomal protein S6 kinase (p70S6K). p70S6K phosphorylates the ribosomal subunit S6 and up-regulates protein synthesis. mTORC2 prevents protein degradation by phosphorylating and inhibiting the forkhead box (FoxO) class of transcription factors. Indeed, FoxO transcription factors induce the expression of atrogin-1/muscle atrophy F-box (MAFbx) and muscle ring finger protein 1 (MuRF-1), two E3 ubiquitin ligases, which promote the ubiquitination and the proteasome-mediated degradation of critical sarcomeric proteins. The ubiquitin-proteasome system mediates muscle atrophy in several conditions, and the oxidative stress plays a key role in the regulation of the proteasome proteolytic activity. Mechanosensory regulation of protein synthesis is determined by high-force contractions that damage the sarcolemma and activate the membrane phospholipid phosphatidic acid, which, in turn, activates mTOR. During resistance exercise, mechanosensory regulation of protein synthesis also involves some transmembrane receptors called focal adhesion kinase (FAK) proteins, which transmit the contractile force through the skeletal muscle architecture and trigger protein synthesis by inducing mTOR activation.
The influence of exercise on muscle mass also involves muscle stem cells. As such, exercise induction of hypertrophy is accompanied by satellite cell fusion to myofibers. Mitochondria are considered as being involved in the regulation of myoblast proliferation/differentiation; therefore, PGC-1α-mediated mitochondrial biogenesis triggered by endurance exercise might possibly influence satellite cell fusion. Interestingly, PGC-1α up-regulation occurs during differentiation. Other signalings triggered by endurance exercise, such as p38 MAPK and Akt, contribute to satellite cell differentiation. Therefore, muscle wasting might be counteracted by endurance training through enhancement of myoblast differentiation and fusion. In addition, it has been suggested that PGC-1α might control muscle wasting pathways. It reduces the FoxO3-associated muscle atrophy, and mice overexpressing PGC-1α are protected from sarcopenia and have an increased lifespan. Moreover, increasing mitochondrial oxidative metabolism and biogenesis protects from atrophy, and this might be achieved by endurance exercise-induced PGC-1α. Regular submaximal aerobic activities have also been found to be beneficial for patients afflicted with Duchenne muscular dystrophy (DMD), while “exercise mimetics” decrease muscle inflammation and inhibit FoxO1 signaling. It has also been shown that exercise inhibits MuRF up-regulation due to diabetes and that this might mediate exercise’s beneficial effects on this disease.
At baseline levels, autophagy is a housekeeping mechanism cleaning cells of aberrant and dysfunctional molecules and organelles, thereby maintaining cell homeostasis. Autophagy is a multi-step process during which a part of the cytoplasm (including intracellular organelles) is sequestered within double-membraned autophagic vacuoles (autophagosomes), which then fuse to lysosomes and become autophagolysosomes (Figure 7). By this mechanism, defective organelles and proteins are digested by lysosomal hydrolases 54. Under stress conditions, autophagy increases and promotes temporary cellular adaptation to unfavorable conditions. It primarily favors survival during nutritional stress imposed by decreased nutrients; the degradation of intracellular material through autophagy becomes an alternative source of energy 55.
Autophagy and skeletal muscle mass maintenance
The role of autophagy in the maintenance of muscle mass is controversial. While excessive autophagy is detrimental to skeletal muscle and contributes to muscle wasting, basal autophagy is required for the maintenance of skeletal muscle homeostasis and integrity 56. The autophagy-lysosome system is activated in several atrophy conditions such as fasting, caloric restriction, cancer cachexia, aging, disuse, and denervation 57. Conversely, the key role of autophagy in skeletal muscle homeostasis maintenance is supported by the fact that muscle-specific ablation of key autophagy proteins such as Atg7 or Atg5 produces myofiber degeneration and muscle weakness 58. On the other hand, the autophagy-lysosome system is activated in several atrophy conditions such as fasting, caloric restriction, cancer cachexia, aging, disuse, and denervation 56. Moreover, the phenotype of some transgenic mice suggests that autophagy may favor muscle atrophy.
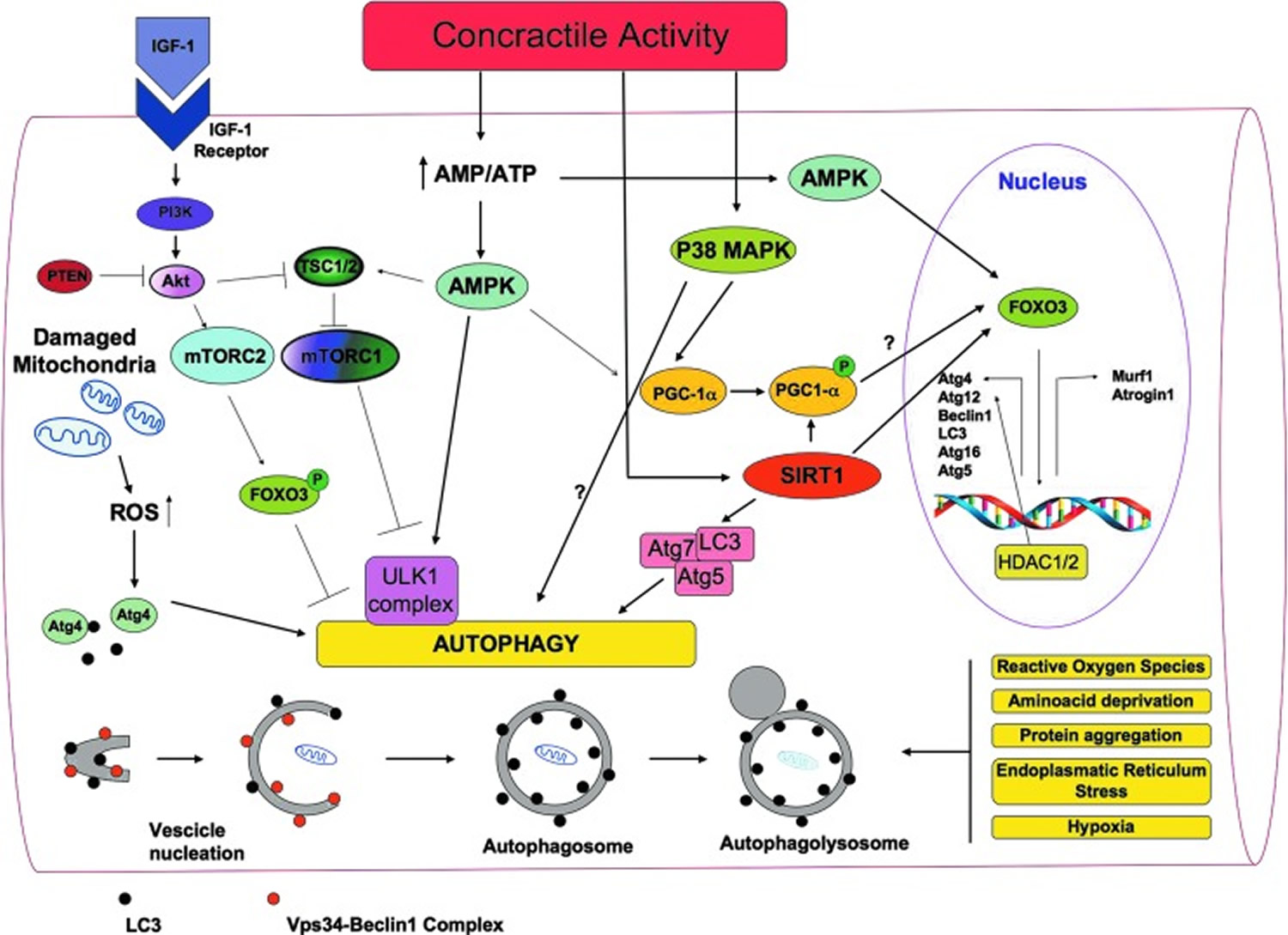
Increasing evidence suggests that exercise triggers autophagy in skeletal muscle and that autophagy mediates some beneficial effects due to exercise. Grumati et al. 59 have revealed an important connection between autophagy and exercise physiology. They have shown that physical training stimulates autophagy in mice skeletal muscles, and that autophagy was able to prevent the accumulation of damaged organelles and to maintain myofiber homeostasis.
It is currently believed that a correct balance between activation and inhibition of autophagy is critical for muscle homeostasis. Too much autophagy causes an excessive removal of crucial cellular components, which leads to muscle atrophy. On the other hand, insufficient autophagy leads to the accumulation of dysfunctional organelles, thus impairing myofiber homeostasis 56.
The involvement of autophagy in exercise-induced remodeling might be related to the two most important functions of autophagy: providing new sources of energy and removing dysfunctional organelles. During exercise, more energy is needed; the requirement of energy generally induces autophagy and it is possible that, as stated earlier, the increase of glucose uptake triggered by exercise depends on autophagy. Moreover, autophagy is the main mechanism for the removal of damaged mitochondria that is necessary to protect myfibers from atrophy. Damaged mitochondria removal is especially needed during exercise when oxidative metabolism and turnover of mitochondria increase. Interestingly, many of the sensors and pathways triggered by exercise in skeletal muscle are involved in the modulation of autophagy.
Figure 7. A general overview of the signaling molecules involved in the regulation of autophagy in skeletal muscles during exercise
What is the best way to build muscle ?
Skeletal muscle tissue is sensitive to the acute and chronic stresses associated with resistance (weight) training. These responses are influenced by the structure of resistance activity (i.e. frequency, load and recovery) as well as the training history of the individuals involved 61. There are histochemical and biochemical data which suggest that resistance (weight) training alters the expression of myosin heavy chains (MyHCs). Specifically, chronic exposure to bodybuilding and power lifting type activity produces shifts towards the MyHC I and IIb isoforms, respectively.
Although many training variables contribute to the performance, cellular and molecular adaptations to resistance exercise, relative intensity (% 1 repetition maximum [%1RM]) appears to be an important factor. In general, relative intensity appears to account for 18-35% of the variance for the muscle hypertrophy response to resistance exercise. On the other hand, fibre type and MyHC transitions were not related to the relative intensity used for training. When competitive lifters were compared, those typically utilising the heaviest loads (> or =90% 1RM), that is weightlifters and powerlifters, exhibited a preferential hypertrophy of type II fibres when compared with body builders who appear to equally hypertrophy both type I and type II fibres. These data suggest that maximal muscle hypertrophy occurs with loads from 80-95% 1RM 52.
Heavy resistance training is associated with increased body weight, lean body mass, and muscle cross-sectional area. The increased muscle cross-sectional area is mainly brought about by hypertrophy of individual muscle fibers 62. There is a greater increase in the area of fast twitch fibers compared to slow twitch fibers. In addition, long-term heavy resistance training may produce fiber proliferation. Mitochondrial volume density decreases in proportion to muscle hypertrophy in response to training. Typically, no capillary neoformation occurs during strength training. Therefore, capillary density decreases consequent to heavy resistance training. It appears, though, that bodybuilders, relying on a high repetition training system, in contrast to Olympic weight- and power lifters, display a small increase in number of capillaries per fiber. Enzyme activities, reflecting oxidative potential; decrease during long-term heavy resistance training, resulting in muscle hypertrophy. Although glycogen storage capacity is enhanced in strength trained athletes, enzyme activities reflecting anaerobic metabolism do not increase in response to heavy resistance exercise.
In a study 63 to investigate the “strength-endurance continuum”, thirty-two untrained men (average age 22.5 years, height 178.3 cm, body mass 77.8 kg) participated in an 8-week progressive resistance-training program.
Subjects were divided into four groups:
- a low repetition group (Low Rep) performing 3-5 repetitions maximum (RM) for four sets of each exercise with 3 min rest between sets and exercises,
- an intermediate repetition group (Int Rep) performing 9-11 repetitions maximum (RM) for three sets with 2 min rest,
- a high repetition group (High Rep) performing 20-28 repetitions maximum (RM) for two sets with 1 min rest, and
- a non-exercising control group (Control).
Three exercises (leg press, squat, and knee extension) were performed 2 days/week for the first 4 weeks and 3 days/week for the final 4 weeks. Maximal strength (one repetition maximum, 1RM), local muscular endurance (maximal number of repetitions performed with 60% of 1RM), and various cardiorespiratory parameters (e.g., maximum oxygen consumption, pulmonary ventilation, maximal aerobic power, time to exhaustion) were assessed at the beginning and end of the study. In addition, pre- and post-training muscle biopsy samples were analyzed for fiber-type composition, cross-sectional area, myosin heavy chain (MyHC) content, and capillarization.
Maximal strength improved significantly more for the Low Rep group compared to the other training groups, and the maximal number of repetitions at 60% 1RM improved the most for the High Rep group. In addition, maximal aerobic power and time to exhaustion significantly increased at the end of the study for only the High Rep group. All three major fiber types (types I, IIA, and IIB) hypertrophied for the Low Rep and Int Rep groups, whereas no significant increases were demonstrated for either the High Rep or Control groups. However, the percentage of type IIB fibers decreased, with a concomitant increase in IIAB fibers for all three resistance-trained groups. These fiber-type conversions were supported by a significant decrease in MyHCIIb accompanied by a significant increase in MyHCIIa. No significant changes in fiber-type composition were found in the control (non-exercise) samples. Although all three training regimens resulted in similar fiber-type transformations (IIB to IIA), the low to intermediate repetition resistance-training programs induced a greater hypertrophic effect compared to the high repetition regimen. The High Rep group, however, appeared better adapted for submaximal, prolonged contractions, with significant increases after training in aerobic power and time to exhaustion. Thus, low and intermediate RM training appears to induce similar muscular adaptations, at least after short-term training in previously untrained subjects. Overall, however, these data demonstrate that both physical performance and the associated physiological adaptations are linked to the intensity and number of repetitions performed, and thus lend support to the “strength-endurance continuum”.
In another study 64 to assess the relationships between human muscle fiber hypertrophy, protein isoform content and maximal Ca2+-activated contractile function following a short-term period of resistance exercise training. Widrick and colleagues 64 put 6 healthy sedentary men (average age 27 yr, height 178 cm, and weight 82.3 kg) under a resistance exercise training program consisting of 36 exercise sessions performed three times per week on nonconsecutive days. The training program used free-weight and machine-based exercises designed to overload the major lower (squats, knee extension, knee flexion, calf raises), upper (bench press, lat pull down, shoulder press, triceps press, biceps curl, seated row), and abdominal muscle groups. During each training session, subjects completed three sets of 5–10 of the exercises listed above (divided approximately equally between those targeting the upper and lower body). Subjects performed 12 repetitions per set during the first 2 wk of the training program. Thereafter, one weekly session was performed at 10 repetitions per set, the second session at 8 repetitions per set, and the third weekly session at 6 repetitions per set. During all sessions, the training resistance was adjusted so that subjects were able to complete only the specified number of repetitions, plus or minus one repetition. This nonlinear periodized program was used to maximize training adaptations. All exercise sessions were supervised by one of the investigators or by a trained assistant.
After 12 weeks of exercise, the volunteers lean body mass rose 4% over the course of the training program (from 63.7 ± 2.8 to 66.4 ± 2.3 kg), whereas total body mass was unchanged. Lower body neuromuscular strength, as assessed by the six-repetition maximum for leg press exercise, rose from a pretraining value of 1,524 ± 99 to 1,791 ± 69 N at the 4th week; 2,241 ± 117 N at the 8th week and 2,532 ± 115 N at the 12th week of training. Over the course of the training program, leg press six-repetition maximum strength increased 62% relative to total body mass (from 18.5 ± 0.8 to 30.0 ± 1.5 N/kg body mass) or 61% relative to lean body mass (from 23.8 ± 0.7 to 38.3 ± 2.1 N/kg lean body mass).
The relative number of fibers containing type IIa MyHC increased from 30% before training to 55% after training, whereas the relative number of single fibers containing type IIa and type IIx MyHC fell from 22 to 3%. Posttraining fibers containing type IIx or type I/IIa MyHC were relatively rare. Consequently, 94% of the pre- and 100% of the posttraining fibers studied contained either type I, type IIa, or type IIa/IIx MyHC 64. The results after twelve weeks of progressive resistance exercise training, sufficient to increase neuromuscular strength by >60%, resulted in significant hypertrophy of fibers containing type I, IIa, or IIa/IIx MyHC. Peak Ca2+-activated force and absolute peak power rose in direct proportion with the increase in fiber cross sectional area, whereas unloaded shortening velocity and power per fiber volume were unaffected by training. These data are consistent with the resistance training-induced increases in slow- and fast-fiber cross-sectional area reported in the histochemical literature 65, 66. It seems likely that the increased neuromuscular power observed after strength training is due, at least in part, to the greater potential of individual muscle fibers to produce power. Although both type I and II fibers hypertrophied, the type II fibers demonstrated a greater capacity for hypertrophy, were more varied in their range of sizes, and were larger than type I fibers both pre- and posttraining. The contribution of the type II fibers would be particularly important in this regard as they produce sixfold greater power than the type I fibers. In conclusion, resistance training resulted in hypertrophy of the total muscle cross sectional area and fiber areas with no change in estimated fiber number, whereas increase in capillary number were proportional to muscle fiber growth.
The effects of diet types (macronutrient composition; eating styles) and their influence on body composition
Diets primarily focused on fat loss are driven by a sustained caloric deficit. The higher the baseline body fat level, the more aggressively the caloric deficit may be imposed 67. However, a slower rates of weight loss can better preserve lean mass in leaner subjects.
Caloric intake
In determining an appropriate caloric intake, it should be noted that the tissue lost during the course of an energy deficit is influenced by the size of the energy deficit. While greater deficits yield faster weight loss, the percentage of weight loss coming from lean body mass (LBM) tends to increase as the size of the deficit increases 68, 69, 70. In studies of weight loss rates, weekly losses of 1 kg compared to 0.5 kg over 4 weeks resulted in a 5% decrease in bench press strength and a 30% greater reduction in testosterone levels in strength training women 71. Weekly weight loss rates of 1.4% of bodyweight compared to 0.7% in athletes during caloric restriction lasting four to eleven weeks resulted in reductions of fat mass of 21% in the faster weight loss group and 31% in the slower loss group. In addition, LBM increased on average by 2.1% in the slower loss group while remaining unchanged in the faster loss group. Worthy of note, small amounts of LBM were lost among leaner subjects in the faster loss group 69.
Therefore, weight loss rates that are more gradual may be superior for LBM retention. At a loss rate of 0.5 kg per week (assuming a majority of weight lost is fat mass), a 70 kg athlete at 13% body fat would need to be no more than 6 kg to 7 kg over their contest weight in order to achieve the lowest body fat percentages recorded in competitive bodybuilders following a traditional three month preparation 72, 73. If a competitor is not this lean at the start of the preparation, faster weight loss will be required which may carry a greater risk for LBM loss.
In a study of bodybuilders during the twelve weeks before competition, male competitors reduced their caloric intake significantly during the latter half and subsequently lost the greatest amount of LBM in the final three weeks 74. Therefore, diets longer than two to four months yielding weight loss of approximately 0.5 to 1% of bodyweight weekly may be superior for LBM retention compared to shorter or more aggressive diets. Ample time should be allotted to lose body fat to avoid an aggressive deficit and the length of preparation should be tailored to the competitor; those leaner dieting for shorter periods than those with higher body fat percentages. It must also be taken into consideration that the leaner the competitor becomes the greater the risk for LBM loss 75, 70. As the availability of adipose tissue declines the likelihood of muscle loss increases, thus it may be best to pursue a more gradual approach to weight loss towards the end of the preparation diet compared to the beginning to avoid LBM loss.
- Diets focused primarily on gaining lean mass are driven by a sustained caloric surplus to facilitate anabolic processes and support increasing resistance-training demands 67. The composition and magnitude of the surplus, as well as training status of the subjects can influence the nature of the gains.
- A wide range of dietary approaches (low-fat to low-carbohydrate/ketogenic diets and all points between) can be similarly effective for improving body composition.
- Bodybuilders typically employ a higher meal frequency in an attempt to optimize fat loss and muscle preservation. However, the majority of chronic experimental studies have failed to show that different meal frequencies have different influences on bodyweight or body composition 76, 77, 78. Despite this limitation, the available research has consistently refuted the popular belief that a grazing pattern (smaller, more frequent meals) raises energy expenditure compared to a gorging pattern (larger, less frequent meals). Disparate feeding patterns ranging from two to seven meals per day have been compared in tightly controlled studies using metabolic chambers, and no significant differences in 24-hour thermogenesis have been detected 79, 80. Along these lines, Stote et al. [113] found that compared to three meals per day, one meal per day caused slightly more weight and fat loss. Curiously, the one meal per day group also showed a slight gain in lean mass, but this could have been due to the inherent error in the use of bioelectrical impedance analysis (BIA) to measure body composition for body composition assessment 81.
- Increasing dietary protein to levels significantly beyond current recommendations for athletic populations may result in improved body composition. The International Society of Sports Nutrition’s original 2007 position stand on protein intake (1.4–2.0 g/kg) 82 has gained further support from subsequent investigations arriving at similar requirements in athletic populations 83, 84, 85, 86, 87, 88.
- Higher protein intakes (2.3–3.1 g/kg lean mass) may be required to maximize muscle retention in lean, resistance-trained subjects under hypocaloric conditions. Emerging research on very high protein intakes (>3 g/kg) has demonstrated that the known thermic, satiating, and lean-mass-preserving effects of dietary protein might be amplified in resistance-training subjects.
- The collective body of intermittent caloric restriction (intermittent fasting) research demonstrates no significant advantage over daily caloric restriction for improving body composition. Time-restricted feeding typically involves a fasting period of 16–20 hours and a feeding period of 4–8 hours daily. Unsurprisingly, significant weight loss occurs, and this includes a reduction in lean mass as well as fat mass 89, 90. An 8-week trial by Tinsley et al. 91 examined the effect of a 20-hour fasting/4-hour feeding protocol (20/4) done 4 days per week on recreationally active, but untrained subjects. No limitations were placed on the amounts and types of food consumed in the 4-hour eating window. A standardized resistance training program was administered 3 days per week. The time-restricted feeding group lost body weight, due to a significantly lower energy intake (667 kcal less on fasting compared to non-fasting days). Cross sectional area of the biceps brachii and rectus femoris increased similarly in both the time-restricted feeding and normal diet group. No significant changes in body composition (via DXA) were seen between groups. Despite a lack of statistical significance, there were notable effect size differences in lean soft tissue (normal diet gained 2.3 kg, while time-restricted feeding lost 0.2 kg). Although both groups increased strength without significant between-group differences, effect sizes were greater in the time-restricted feeding group for bench press endurance, hip sled endurance, and maximal hip sled strength. This finding should be viewed cautiously given the potential for greater and more variable neurological gains in untrained subjects. A subsequent study by Moro et al. 92 found that in resistance-trained subjects on a standardized training protocol, a 16-hour fasting/8-hour feeding cycle (16/8) resulted in significantly greater fat loss in time-restricted feeding vs. normal diet control group (ND) (1.62 vs. 0.31 kg), with no significant changes in lean mass in either group. Time-restricted feeding’s meals were consumed at 1 pm, 4 pm, and 8 pm. Normal diet’s meals were consumed at 8 am, 1 pm, and 8 pm. Macronutrient intake between the time-restricted feeding and normal diet groups was matched, unlike the aforementioned Tinsley et al. study 91 whereby protein intake was disparate and sub-optimal (1.0 g/kg in the time-restricted feeding group and 1.4 g/kg in the normal diet control group). Subjects in the present study’s time-restricted feeding and normal diet group consumed 1.93 and 1.89 g/kg, respectively. The mechanisms underlying these results are not clear. The authors speculated that increased adiponectin levels in the time-restricted feeding group could have stimulated mitochondrial biogenesis via interacting with PPAR-gamma, in addition to adiponectin acting centrally to increase energy expenditure. However, the time-restricted feeding group also experienced unfavorable changes such as decreased testosterone and triiodothyronine levels.
- Seimon et al. 93 recently published the largest systematic review of intermittent fasting research to date, comparing the effects of intermittent energy restriction (IER) to continuous energy restriction (CER) on body weight, body composition, and other clinical parameters. Their review included 40 studies in total, 12 of which directly compared an intermittent energy restriction (IER) with a continuous energy restriction (CER) condition. They found that overall, the two diet types resulted in “apparently equivalent outcomes” in terms of body weight reduction and body composition change. Interestingly, intermittent energy restriction (IER) was found to be superior at suppressing hunger. The authors speculated that this might be attributable to ketone production in the fasting phases. However, this effect was immaterial since on the whole, intermittent fasting failed to result in superior improvements in body composition or greater weight loss compared to continuous energy restriction (CER). Table 1 outlines the characteristics of the major diet archetypes.
- Dehydration: In an attempt to enhance muscle size and definition by reducing extracellular water content, many bodybuilders engage in fluid, electrolyte, and carbohydrate manipulation in the final days and hours before competing 94, 95. The effect of electrolyte manipulation and dehydration on visual appearance has not been studied, however it may be a dangerous practice 96. Furthermore, dehydration could plausibly degrade appearance considering that extracellular water is not only present in the subcutaneous layer. A significant amount is located in the vascular system. Thus, the common practice of “pumping up” to increase muscle size and definition by increasing blood flow to the muscle with light, repetitive weight lifting prior to stepping on stage 97 could be compromised by dehydration or electrolyte imbalance. Furthermore, dehydration reduces total body hydration. A large percentage of muscle tissue mass is water and dehydration results in decreases in muscle water content 98 and therefore muscle size, which may negatively impact the appearance of muscularity. At this time it is unknown whether dehydration or electrolyte manipulation improves physique appearance. What is known is that these practices are dangerous and have the potential to worsen it. It is unclear if carbohydrate loading has an impact on appearance and if so, how significant the effect is. However, the recommended muscle-sparing practice by some researchers to increase the carbohydrate content of the diet in the final weeks of preparation 99 might achieve any proposed theoretical benefits of carbohydrate loading. If carbohydrate loading is utilized, a trial run before competition once the competitor has reached or nearly reached competition leanness should be attempted to develop an individualized strategy. However, a week spent on a trial run consuming increased carbohydrates and calories may slow fat loss, thus ample time in the diet would be required.
- Carbohydrate Loading: In the final days before competing, bodybuilders commonly practice carbohydrate loading similar to endurance athletes in an attempt to raise muscle-glycogen levels and increase muscle size 100, 101, 102, 97. In the only direct study of this practice, no significant quantitative change in muscle girth was found to occur [208]. However, an isocaloric diet was used, with only a change in the percentage of carbohydrate contributing to the diet. If total calories had also been increased, greater levels of glycogen might have been stored which could have changed the outcome of this study. Additionally, unlike the subjects in this study bodybuilders prior to carbohydrate loading have reduced glycogen levels from a long calorically restricted diet and it is possible in this state that carbohydrate loading might effect a visual change. Furthermore, bodybuilding performance is measured subjectively, thus analysis of girth alone may not discern subtle visual changes which impact competitive success. Lastly, some bodybuilders alter the amount of carbohydrate loaded based on the visual outcome, increasing the amount if the desired visual change does not occur 102. Thus, an analysis of a static carbohydrate load may not accurately represent the dynamic nature of actual carbohydrate loading practices.In fact, in an observational study of competitive bodybuilders in the days before competition who loaded carbohydrates, subjects showed a 4.9% increase in biceps thickness the final day before competition compared to six weeks prior 100. Although it is unknown if this was caused by increased muscle glycogen, it is unlikely it was due to muscle mass accrual since the final weeks of preparation are often marked by decreases not increases in lean mass 99. Future studies of this practice should include a qualitative analysis of visual changes and analyze the effects of concurrent increases in percentage of carbohydrates as well as total calories.
Table 1. Diet categories
| Diet | Composition | Strengths | Limitations |
|---|---|---|---|
| Low-energy diets (LED) | LED: 800–1200 kcal/day VLED: 400–800 kcal/day | Rapid weight loss (1.0–2.5 kg/week, diets involve premade products that eliminate or minimize the need for cooking and planning. | VLED have a higher risk for more severe side-effects, but do not necessary outperform LED in the long-term |
| Low-fat diets (LFD) | LFD: 25–30% fat VLFD: 10–20% fat | LFD have the support of the major health organizations due to their large evidence basis in the literature on health effects. Flexible macronutrient range. Does not indiscriminately vilify foods based on CHO content. | Upper limits of fat allowance may falsely convey the message that dietary fat is inherently antagonistic to body fat reduction. VLFD have a scarce evidence basis in terms of comparative effects on body composition, and extremes can challenge adherence. |
| Low-carbohydrate diets (LCD) | 50–150 g CHO, or up to 40% of kcals from CHO | Defaults to higher protein intake. Large amount of flexibility in macronutrient proportion, and by extension, flexibility in food choices. Does not indiscriminately prohibit foods based on fat content. | Upper limits of CHO allowance may falsely convey the message that CHO is inherently antagonistic to body fat reduction. |
| Ketogenic diets (KD) | Maximum of ~50 g CHO Maximum of ~10% CHO | Defaults to higher protein intake. Suppresses appetite/controls hunger, causes spontaneous reductions in kcal intake under non-calorically restricted conditions. Simplifies the diet planning and decision-making process. | Excludes/minimizes high-CHO foods which can be nutrient dense and disease-preventive. Can compromise high-intensity training output. Has not shown superior effects on body composition compared to non-KD when protein and kcals are matched. Dietary extremes can challenge long-term adherence. |
| High-protein diets (HPD) | HPD: ≥ 25% of total kcals, or 1.2–1.6 g/kg (or more) Super HPD: > 3 g/kg | HPD have a substantial evidence basis for improving body composition compared to RDA levels (0.8 g/kg), especially when combined with training. Super-HPD have an emerging evidence basis for use in trained subjects seeking to maximize intake with minimal-to-positive impacts on body composition. | May cause spontaneous reductions in total energy intake that can antagonize the goal of weight gain. Potentially an economical challenge, depending on the sources. High protein intakes could potentially displace intake of other macronutrients, leading to sub-optimal intakes (especially CHO) for athletic performance goals. |
| Intermittent fasting (IF) | Alternate-day fasting (ADF): alternating 24-h fast, 24-h feed. Whole-day fasting (WDF): 1–2 complete days of fasting per week. Time-restricted feeding (TRF): 16–20-h fast, 4–8-h feed, daily. | ADF, WDF, and TRF have a relatively strong evidence basis for performing equally and sometimes outperforming daily caloric restriction for improving body composition. ADF and WDF have ad libitum feeding cycles and thus do not involve precise tracking of intake. TRF combined with training has an emerging evidence basis for the fat loss while maintaining strength. | Questions remain about whether IF could outperform daily linear or evenly distributed intakes for the goal of maximizing muscle strength and hypertrophy. IF warrants caution and careful planning in programs that require optimal athletic performance. |
- The long-term success of a diet depends upon compliance and suppression or circumvention of mitigating factors such as adaptive thermogenesis. Joosen and Westerterp 104 examined the literature (11 studies) to see if “adaptive thermogenesis” existed in overeating experiments. No evidence beyond the theoretical costs of increased body size and thermic effect of food were found. Nevertheless, there is substantial interindividual variability in the energetic response to overfeeding. They found in overfeeding experiments, weight gain is often less than expected from the energy excess intake. In part this is the result of an obligatory increase in energy expenditure associated with the increased body weight and lean mass 105 and the larger amount of food to be digested and absorbed 106. However, evidence for adaptive thermogenesis as a mechanism to explain interindividual differences in weight gain on the same overeating regimen is still inconsistent and hard to prove 107
- There is a paucity of research on women and older populations, as well as a wide range of untapped permutations of feeding frequency and macronutrient distribution at various energetic balances combined with training. Behavioral and lifestyle modification strategies are still poorly researched areas of weight management.
Role of Protein and Amino Acids in promoting Lean Mass gain with resistance exercise
Amino acids are major nutrient regulators of muscle protein turnover. After protein ingestion, hyperaminoacidemia stimulates increased rates of skeletal muscle protein synthesis, suppresses muscle protein breakdown and promotes net muscle protein gain for several hours 108. These acute observations form the basis for strategized protein intake to promote lean mass gain, or prevent lean mass loss over the long term. However, factors such as protein dose, protein source, and timing of intake are important in mediating the anabolic effects of amino acids on skeletal muscle and must be considered within the context of evaluating the reported efficacy of long-term studies investigating protein supplementation as part of a dietary strategy to promote lean mass accretion and/or prevent lean mass loss. Current research suggests that dietary protein supplementation can augment resistance exercise-mediated gains in skeletal muscle mass and strength and can preserve skeletal muscle mass during periods of diet-induced energy restriction 108. Perhaps less appreciated, protein supplementation can augment resistance training-mediated gains in skeletal muscle mass even in individuals habitually consuming ‘adequate’ (i.e., >0.8 g kg/day) protein. Additionally, overfeeding energy with moderate to high-protein intake (15–25 % protein or 1.8–3.0 g kg/day) is associated with lean, but not fat mass accretion, when compared to overfeeding energy with low protein intake (5 % protein or ~0.68 g kg/day) 108. Amino acids represent primary nutrient regulators of skeletal muscle anabolism, capable of enhancing lean mass accretion with resistance exercise and attenuating the loss of lean mass during periods of energy deficit, although factors such as protein dose, protein source, and timing of intake are likely important in mediating these effects.
Adequate Protein Consumption
In a review by Phillips and Van Loon [28], it is suggested that a protein intake of 1.8-2.7 g/kg for athletes training in hypocaloric conditions may be optimal. While this is one of the only recommendations existing that targets athletes during caloric restriction, this recommendation is not given with consideration to bodybuilders performing concurrent endurance and resistance training at very low levels of body fat. The collective agreement among reviewers is that a protein intake of 1.2-2.2 g/kg is sufficient to allow adaptation to training for athletes whom are at or above their energy needs 109, 110. However, bodybuilders during their contest preparation period typically perform resistance and cardiovascular training, restrict calories and achieve very lean conditions 111, 74. Each of these factors increases protein requirements and when compounded may further increase protein needs 112. Therefore, optimal protein intakes for bodybuilders during contest preparation may be significantly higher than existing recommendations. However, the recently published systematic review by Helms et al. 112 on protein intakes in resistance-trained, lean athletes during caloric restriction suggests a range of 2.3-3.1 g/kg of LBM, which may be more appropriate for bodybuilding. Moreover, the authors suggest that the lower the body fat of the individual, the greater the imposed caloric deficit and when the primary goal is to retain LBM, the higher the protein intake (within the range of 2.3-3.1 g/kg of LBM) should be.
Carbohydrate Intake
While it is true that resistance training utilizes glycogen as its main fuel source 113, total caloric expenditure of strength athletes is less than that of mixed sport and endurance athletes. Thus, authors of a recent review recommend that carbohydrate intakes for strength sports, including bodybuilding, be between 4–7 g/kg depending on the phase of training 114. However, in the specific case of a bodybuilder in contest preparation, achieving the necessary caloric deficit while consuming adequate protein and fat would likely not allow consumption at the higher end of this recommendation.
Satiety and fat loss generally improve with lower carbohydrate diets; specifically with higher protein to carbohydrate ratios 115, 116. In terms of performance and health, low carbohydrate diets are not necessarily as detrimental as typically espoused 117. In a recent review, it was recommended for strength athletes training in a calorically restricted state to reduce carbohydrate content while increasing protein to maximize fat oxidation and preserve LBM 109. However, the optimal reduction of carbohydrate and point at which carbohydrate reduction becomes detrimental likely needs to be determined individually.
While it appears low carbohydrate, high protein diets can be effective for weight loss, a practical carbohydrate threshold appears to exist where further reductions negatively impact performance and put one at risk for LBM losses. In support of this notion, researchers studying bodybuilders during the final 11 weeks of contest preparation concluded that had they increased carbohydrate during the final weeks of their diet they may have mitigated metabolic and hormonal adaptations that were associated with reductions in LBM 118.
Therefore, once a competitor has reached or has nearly reached the desired level of leanness, it may be a viable strategy to reduce the caloric deficit by an increase in carbohydrate. For example, if a competitor has reached competition body fat levels (lacking any visible subcutaneous fat) and is losing half a kilogram per week (approximately a 500 kcals caloric deficit), carbohydrate could be increased by 25-50 g, thereby reducing the caloric deficit by 100-200 kcals in an effort to maintain performance and LBM. However, it should be noted that like losses of LBM, decrements in performance may not affect the competitive outcome for a bodybuilder. It is possible that competitors who reach the leanest condition may experience unavoidable drops in performance.
Fat Intake
Body composition and caloric restriction may play greater roles in influencing testosterone levels that fat intake. During starvation, a reduction in testosterone occurs in normal weight, but not obese, males 119. In addition, rate of weight loss may influence testosterone levels. Weekly target weight loss rates of 1 kg resulted in a 30% reduction in testosterone compared to target weight loss rates of 0.5 kg per week in resistance trained women of normal weight 120.
Reductions in the percentage of dietary fat in isocaloric diets from approximately 40% to 20% has resulted in modest, but significant, reductions in testosterone levels 121. However, distinguishing the effects of reducing total dietary fat on hormonal levels from changes in caloric intake and percentages of saturated and unsaturated fatty acids in the diet is difficult 122. However, a drop in testosterone does not equate to a reduction in LBM. In direct studies of resistance trained athletes undergoing calorically restricted high protein diets, low fat interventions that maintain carbohydrate levels appear to be more effective at preventing LBM loses than lower carbohydrate, higher fat approaches. These results might indicate that attempting to maintain resistance training performance with higher carbohydrate intakes is more effective for LBM retention than attempting to maintain testosterone levels with higher fat intakes.
While cogent arguments for fat intakes between 20 to 30% of calories have been made to optimize testosterone levels in strength athletes 123, in some cases this intake may be unrealistic in the context of caloric restriction without compromising sufficient protein or carbohydrate intakes. While dieting, low carbohydrate diets may degrade performance 124 and lead to lowered insulin and IGF-1 which appear to be more closely correlated to LBM preservation than testosterone 118. Thus, a lower end fat intake between 15-20% of calories, which has been previously recommended for bodybuilders 125, can be deemed appropriate if higher percentages would reduce carbohydrate or protein below ideal ranges.
Table 2. Dietary recommendations for bodybuilding contest preparation
| Diet component | Recommendation |
|---|---|
| Protein (g/kg of LBM) | 2.3-3.1 |
| Fat (% of total calories) | 15-30% |
| Carbohydrate (% of total calories) | remaining |
| Weekly weight loss (% of body weight) | 0.5-1% |
Note: It must be noted that there is a high degree of variability in the way that individuals respond to diets. If training performance degrades it may prove beneficial to decrease the percentage of calories from dietary fat within these ranges in favor of a greater proportion of carbohydrate. Finally, while outside of the norm, some competitors may find that they respond better to diets that are higher in fat and lower in carbohydrate than recommended in this review. Therefore, monitoring of individual response over a competitive career is suggested. There is no evidence of any relationships with bone structure or regional subcutaneous fat distribution with any response to specific macronutrient ratios in bodybuilders or athletic populations. Bodybuilders, like others athletes, most likely operate best on balanced macronutrient intakes tailored to the energy demands of their sport 126. While the majority of competitors will respond best to the fat and carbohydrate guidelines proposed, the occasional competitor will undoubtedly respond better to a diet that falls outside of these suggested ranges. Careful monitoring over the course of a competitive career is required to determine the optimal macronutrient ratio for pre-contest dieting.
Timing and consumption of protein and/or carbohydrate during workouts
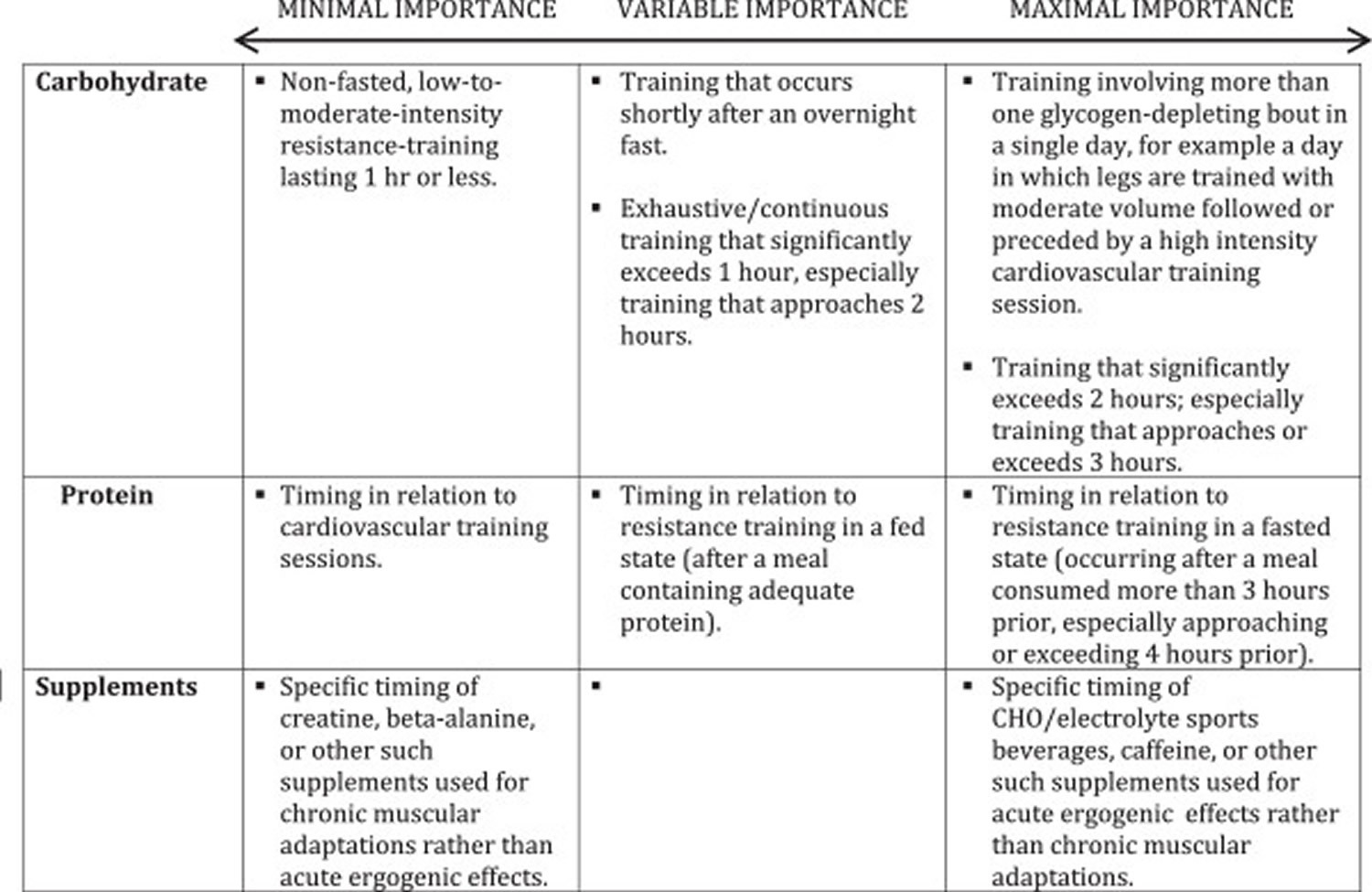
Questions remain about the utility of consuming protein and/or carbohydrate during bodybuilding-oriented training bouts. Since these bouts typically do not resemble endurance bouts lasting 2 hours or more, nutrient consumption during training is not likely to yield any additional performance-enhancing or muscle -sparing benefits if proper pre-workout nutrition is in place. In the exceptional case of resistance training sessions that approach or exceed two hours of exhaustive, continuous work, it might be prudent to employ tactics that maximize endurance capacity while minimizing muscle damage. This would involve approximately 8–15 g protein co-ingested with 30–60 g carbohydrate in a 6-8% solution per hour of training 127. Nutrient timing is an intriguing area of study that focuses on what might clinch the competitive edge. In terms of practical application to resistance training bouts of typical length, Aragon and Schoenfeld 128 recently suggested a protein dose corresponding with 0.4-0.5 g/kg bodyweight consumed at both the pre- and post-exercise periods. However, for objectives relevant to bodybuilding, the current evidence indicates that the global macronutrient composition of the diet is likely the most important nutritional variable related to chronic training adaptations. Table 3 below provides a continuum of importance with bodybuilding-specific context for nutrient timing.
Table 3. Continuum of nutrient & supplement timing importance
Nutritional Supplements
Some bodybuilders and athletes turn to dietary supplements to help them increase muscle size and definition 129. However, many bodybuilding products marketed as dietary supplements have been found to contain other ingredients that can be harmful. Use caution and talk with your health care provider before you begin taking any supplement to gain strength or muscle size. Furthermore, natural bodybuilding federations have extensive banned substance lists 130. It should be noted that there are considerably more supplements that are used by bodybuilders and sold on the market. However, an exhaustive review of all of the supplements commonly used by bodybuilders that often lack supporting data is beyond the scope of this article.
- Multivitamin and mineral supplements are unnecessary for athletes or other physically active people who eat a well-balanced diet and enough calories. The safety of supplements used for bodybuilding remains an issue of concern (see Safety below).
- There is no scientific evidence that other dietary supplements, such as choline, methoxyisoflavone, zinc/magnesium aspartate, nitric oxide precursors, and chromium, are effective for building strength and muscle mass.
- Evidence suggests that creatine, a popular dietary supplement, may enhance the effects of vigorous exercise on strength, muscle mass, and endurance, but it may also cause fluid weight gain, nausea, cramping, and diarrhea.
Safety
- Many bodybuilding products marketed as dietary supplements have been found to be deceptively labeled and to contain hidden ingredients that can be harmful, such as anabolic steroids, compounds chemically similar to them, or other substances that don’t qualify as dietary ingredients.
- In April 2013, the U.S. Food and Drug Administration issued a warning to consumers to avoid products containing the stimulant dimethylamylamine (DMAA). DMAA can elevate blood pressure and lead to other problems, such as a heart attack.
- Evidence suggests that creatine (an amino acid produced by the body) supplements may be safe for short-term use in healthy adults, but the American College of Sports Medicine recommends against anyone younger than age 18 using it to enhance athletic performance.
- Some dietary supplements may have side effects and some may interact with drugs or other supplements. Some vitamins and minerals are harmful at high doses. Talk with your health care provider before using a dietary supplement to increase muscle size and strength.
Micronutrients
Several previous studies have observed deficiencies in intakes of micronutrients, such as vitamin D, calcium, zinc, magnesium, and iron, in dieting bodybuilders 131, 132. However, it should be noted that these studies were all published nearly 2 decades ago and that micronutrient deficiencies likely occurred due to elimination of foods or food groups and monotony of food selection 133, 132. Therefore, future studies are needed to determine if these deficiencies would present while eating a variety of foods and using the contest preparation approach described herein. Although the current prevalence of micronutrient deficiencies in competitive bodybuilders is unknown, based on the previous literature, a low-dose micronutrient supplement may be beneficial for natural bodybuilders during contest preparation; however, future studies are needed to verify this recommendation.
Caffeine
Caffeine is perhaps the most common pre-workout stimulant consumed by bodybuilders. Numerous studies support the use of caffeine to improve performance during endurance training 134, 135, sprinting 136, 137, and strength training 138, 139, 140. However, not all studies support use of caffeine to improve performance in strength training 141, 142. It should be noted that many of the studies that found increases in strength training performance supplemented with larger (5–6 mg/kg) dosages of caffeine. However, this dosage of caffeine is at the end of dosages that are considered safe (6 mg/kg/day) 143. Additionally, it appears that regular consumption of caffeine may result in a reduction of ergogenic effects 144. Therefore, it appears that 5–6 mg/kg caffeine taken prior to exercise is effective in improving exercise performance; however, caffeine use may need to be cycled in order for athletes to obtain the maximum performance enhancing effect.
Beta-alanine
Beta-alanine (BA) is becoming an increasingly popular supplement among bodybuilders. Once consumed, BA enters the circulation and is up-taken by skeletal muscle where it is used to synthesize carnosine, a pH buffer in muscle that is particularly important during anaerobic exercise such as sprinting or weightlifting 145. Indeed, consumption of 6.4 g BA daily for four weeks has been shown to increase muscle carnosine levels by 64.2% 146. Moreover, supplementation with BA for 4–10 weeks has been shown to increase knee extension torque by up to 6%, improve workload and time to fatigue during high intensity cardio, improve muscle resistance to fatigue during strength training, increase lean mass by approximately 1 kg and significantly reduce perceptions of fatigue. Additionally, the combination of BA and CM may increase performance of high intensity endurance exercise and has been shown to increase lean mass and decrease body fat percentage more than CM alone. However, not all studies have shown improvements in performance with BA supplementation. To clarify these discrepancies, Hobson et al. 147 conducted a meta-analysis of 15 studies on BA supplementation and concluded that BA significantly increased exercise capacity and improved exercise performance on 60-240 seconds and >240 seconds exercise bouts.
Although BA appears to improve exercise performance, the long-term safety of BA has only been partially explored. Currently, the only known side effect of BA is unpleasant symptoms of parasthesia reported after consumption of large dosages; however, this can be minimized through consumption of smaller dosages throughout the day 146. While BA appears to be relatively safe in the short-term, the long-term safety is unknown. In cats, an addition of 5 percent BA to drinking water for 20 weeks has been shown to deplete taurine and result in damage to the brain; however, taurine is an essential amino acid for cats but not for humans and it is unknown if the smaller dosages consumed by humans could result in similar effects 148. BA may increase exercise performance and increase lean mass in bodybuilders and currently appears to be safe; however, studies are needed to determine the long-term safety of BA consumption.
Beta-hydroxy-beta-methylbutyrate
Beta-hydroxy-beta-methylbutyrate (HMB) is a metabolite of the amino acid leucine that has been shown to decrease muscle protein catabolism and increase muscle protein synthesis 149, 150. The safety of HMB supplementation has been widely studied and no adverse effects on liver enzymes, kidney function, cholesterol, white blood cells, hemoglobin, or blood glucose have been observed 151, 152. Furthermore, two meta-analyses on HMB supplementation have concluded that HMB is safe and does not result in any major side effects 151. HMB may actually decrease blood pressure, total and LDL cholesterol, especially in hypercholesterolemic individuals.
HMB is particularly effective in catabolic populations such as the elderly and patients with chronic disease 153. However, studies on the effectiveness of HMB in trained, non-calorically restricted populations have been mixed. Reasons for discrepancies in the results of HMB supplementation studies in healthy populations may be due to many factors including clustering of data in these meta-analysis to include many studies from similar groups, poorly designed, non-periodized training protocols, small sample sizes, and lack of specificity between training and testing conditions 154. However, as a whole HMB appears to be effective in a majority of studies with longer-duration, more intense, periodized training protocols and may be beneficial to bodybuilders, particularly during planned over-reaching phases of training 155. While the authors hypothesize that HMB may be effective in periods of increased catabolism, such as during contest preparation, the efficacy of HMB on maintenance of lean mass in dieting athletes has not been investigated in a long-term study. Therefore, future studies are needed to determine the effectiveness of HMB during caloric restriction in healthy, lean, trained athletes.
Creatine
Creatine is a naturally-occurring amino acid (protein building block) that’s found in meat and fish, and also made by the human body predominately in the liver, kidneys, and to a lesser extent in the pancreas 156. Creatine is produced in your body at an amount of about 1 g/day 157. The remainder of the creatine available to the body is obtained through the diet at about 1 g/day for an omnivorous diet 157. 95% of the bodies creatine stores are found in the skeletal muscle and the remaining 5% is distributed in the brain, liver, kidney, and testes 158.
Creatine is chemically known as a non-protein nitrogen; a compound which contains nitrogen but is not a protein per se 159. It is synthesized in the liver and pancreas from the amino acids arginine, glycine, and methionine 159, 160. Approximately 95% of the body’s creatine is stored in skeletal muscle. Additionally, small amounts of creatine are also found in the brain and testes 161, 162. About two thirds of the creatine found in skeletal muscle is stored as phosphocreatine (PCr) while the remaining amount of creatine is stored as free creatine 161. The total creatine pool (PCr + free creatine) in skeletal muscle averages about 120-140 grams for a 70 kg individual 163, depending on the skeletal muscle fiber type 158 and quantity of muscle mass 164. However, the average human has the capacity to store up to 160 grams of creatine under certain conditions 165, 166. The body breaks down about 1 – 2% of the creatine pool per day (about 1–2 grams/day) into creatinine in the skeletal muscle 159. The body production and dietary intake matches the rate of creatinine production from the degradation of phosphocreatine and creatine at 2.6% and 1.1%/d respectively. In general, oral creatine supplementation leads to an increase of creatine levels within the body. Creatine can be cleared from the blood by saturation into various organs and cells or by renal filtration then excreted in urine 158.
Creatine stores can be replenished by obtaining creatine in the diet or through endogenous synthesis of creatine from glycine, arginine, and methionine 167, 168. Dietary sources of creatine include meats and fish. Large amounts of fish and meat must be consumed in order to obtain gram quantities of creatine. Whereas dietary supplementation of creatine provides an inexpensive and efficient means of increasing dietary availability of creatine without excessive fat and/or protein intake.
Three amino acids (glycine, arginine and methionine) and three enzymes (L-arginine:glycine amidinotransferase, guanidinoacetate methyltransferase and methionine adenosyltransferase) are required for creatine synthesis. The impact creatine synthesis has on glycine metabolism in adults is low, however the demand is more appreciable on the metabolism of arginine and methionine 164.
As creatine is predominately present in the diet from meats, vegetarians have lower resting creatine concentrations 169. It is converted into creatine phosphate or phosphocreatine and stored in the muscles, where it is used for energy 156. During high-intensity, short-duration exercise, such as lifting weights or sprinting, phosphocreatine is converted into ATP, a major source of energy within the human body.
Biochemically speaking, the energy supplied to rephosphorylate adenosine diphosphate (ADP) to adenosine triphosphate (ATP) during and following intense exercise is largely dependent on the amount of phosphocreatine (PCr) stored in the muscle 170, 165. As phosphocreatine stores become depleted during intense exercise, energy availability diminishes due to the inability to resynthesize ATP at the rate required to sustained high-intensity exercise 170, 165. Consequently, the ability to maintain maximal-effort exercise declines. The availability of phosphocreatine in the muscle may significantly influence the amount of energy generated during brief periods of high-intensity exercise. Furthermore, it has been hypothesized that increasing muscle creatine content, via creatine supplementation, may increase the availability of phosphocreatine allowing for an accelerated rate of resynthesis of ATP during and following high-intensity, short-duration exercise 170, 165. Theoretically, creatine supplementation during training may lead to greater training adaptations due to an enhanced quality and volume of work performed. In terms of potential medical applications, creatine is intimately involved in a number of metabolic pathways. For this reason, medical researchers have been investigating the potential therapeutic role of creatine supplementation in a variety of patient populations.
Creatine supplements are popular among body builders and competitive athletes. As an oral supplement, the most widely used and researched form is creatine monohydrate 157. When orally ingested, creatine monohydrate has shown to improve exercise performance and increase fat free mass 171, 172, 173, 174, 175. The attraction of creatine is that it may increase lean muscle mass and enhance athletic performance, particularly during high-intensity, short-duration sports (like high jumping and weight lifting).
However, not all human studies show that creatine improves athletic performance 156. Nor does every person seem to respond the same way to creatine supplements. For example, people who tend to have naturally high stores of creatine in their muscles don’t get an energy-boosting effect from extra creatine. Preliminary clinical studies also suggest that creatine’s ability to increase muscle mass and strength may help fight muscle weakness associated with illnesses, such as heart failure and muscular dystrophy.
Creatine supplementation Responders vs. Non-responders
Syrotuik and Bell 176 investigated the physical characteristics of responder and non-responder subjects to creatine supplementation in recreationally resistance trained men with no history of creatine monohydrate usage. The supplement group was asked to ingest a loading dosage of 0.3 g/kg/d for 5 days. The physiological characteristics of responders were classified using Greenhaff et al 177 criterion of >20 mmol/kg dry weight increase in total intramuscular creatine and phosphocreatine and non responders as <10 mmol/kg dry weight increase, a third group labeled quasi responders were also used to classify participants who fell in between the previously mentioned groups (10-20 mmol/kg dry weight). Overall, the supplemented group showed a mean increase in total resting muscle creatine and phosphocreatine of 14.5% (from 111.12 ± 8.87 mmol/kg dry weight to 127.30 ± 9.69 mmol/kg dry weight) whilst the placebo group remained relatively unaffected (from 115.70 ± 14.99 mmol/kg dry weight to 111.74 ± 12.95 mmol/kg dry weight). However when looking at individual cases from the creatine group the results showed a variance in response. From the 11 males in the supplemented group, 3 participants were responders (mean increase of 29.5 mmol/kg dry weight or 27%), 5 quasi responders (mean increase of 14.9 mmol/kg dry weight or 13.6%) and 3 non-responders (mean increase of 5.1 mmol/kg dry weight or 4.8%). Using muscle biopsies of the vastus lateralis, a descending trend for groups and mean percentage fiber type was observed. Responders showed the greatest percentage of type II fibers followed by quasi responders and non-responders. The responder and quasi responder groups had an initial larger cross sectional area for type I, type IIa and type IIx fibers. The responder group also had the greatest mean increase in the cross sectional area of all the muscle fiber types measured (type I, type IIa and type IIx increased 320, 971 and 840 μm2 respectively) and non-responders the least (type I, type IIa and type IIx increased 60, 46 and 78 μm2 respectively). There was evidence of a descending trend for responders to have the highest percentage of type II fibers; furthermore, responders and quasi responders possessed the largest initial cross sectional area of type I, IIa and IIx fibers. Responders were seen to have the lowest initial levels of creatine and phosphocreatine. This has also been observed in a previous study 178 which found that subjects whose creatine levels were around 150 mmol/Kg dry mass did not have any increments in their creatine saturation due to creatine supplementation, neither did they experience any increases of creatine uptake, phosphocreatine resynthesis and performance. This would indicate a limit maximum size of the creatine pool.
In summary responders are those individuals with a lower initial level of total muscle creatine content, greater population of type II fibers and possess higher potential to improve performance in response to creatine supplementation.
Effects of creatine supplementation on Muscle Mass
Cribb et al (2007) 179 observed greater improvements on 1RM, lean body mass, fiber cross sectional area and contractile protein in trained young males when resistance training was combined with a multi-nutrient supplement containing 0.1 g/kg/d of creatine, 1.5 g/kg/d of protein and carbohydrate compared with protein alone or a protein carbohydrate supplement without the creatine. These findings were novel because at the time no other research had noted such improvements in body composition at the cellular and sub cellular level in resistance trained participants supplementing with creatine. The amount of creatine consumed in the study by Cribb et al 179 was greater than the amount typically reported in previous studies (a loading dose of around 20 g/d followed by a maintenance dose of 3-5 g/d is generally equivalent to approximately 0.3 g/kg/d and 0.03 g/kg/d respectively) and the length of the supplementation period or absence of resistance exercise may explain the observed transcriptional level changes that were absent in previous studies 180, 181.
Deldicque et al 182 found a 250%, 45% and 70% increase for collagen mRNA, glucose transporter 4 (GLUT4) and Myosin heavy chain IIA, respectively after 5 days creatine loading protocol (21 g/d). The authors speculated that creatine in addition to a single bout of resistance training can favor an anabolic environment by inducing changes in gene expression after only 5 days of supplementation.
When creatine supplementation is combined with heavy resistance training, muscle insulin like growth factor (IGF-1) concentration has been shown to increase. Burke et al 183 examined the effects of an 8 week heavy resistance training protocol combined with a 7 day creatine loading protocol (0.25 g/d/kg lean body mass) followed by a 49 day maintenance phase (0.06 g/kg lean mass) in a group of vegetarian and non-vegetarian, novice, resistance trained men and women. Compared to placebo, creatine groups produced greater increments in IGF-1 (78% Vs 55%) and body mass (2.2 Vs 0.6 kg). Additionally, vegetarians within the supplemented group had the largest increase of lean mass compared to non vegetarian (2.4 and 1.9 kg respectively). Changes in lean mass were positively correlated to the modifications in intramuscular total creatine stores which were also correlated with the modified levels of intramuscular IGF-1. The authors suggested that the rise in muscle IGF-1 content in the creatine group could be due to the higher metabolic demand created by a more intensely performed training session. These amplifying effects could be caused by the increased total creatine store in working muscles. Even though vegetarians had a greater increase in high energy phosphate content, the IGF-1 levels were similar to the amount observed in the non vegetarian groups. These findings do not support the observed correlation pattern by which a low essential amino acid content of a typical vegetarian diet should reduce IGF-1 production 184. According to authors opinions it is possible that the addition of creatine and subsequent increase in total creatine and phosphocreatine storage might have directly or indirectly stimulated production of muscle IGF-I and muscle protein synthesis, leading to an increased muscle hypertrophy 183.
Effects of creatine ingestion to improve recovery from injury, muscle damage and oxidative stress induced by exercise
Creatine supplementation may also be of benefit to injured athletes. Op’t Eijnde et al 185 noted that the expected decline in GLUT4 content after being observed during a immobilization period can be offset by a common loading creatine (20g/d) supplementation protocol. In addition, combining creatine monohydrate 15g/d for 3 weeks following 5 g/d for the following 7 weeks positively enhances GLUT4 content, glycogen, and total muscle creatine storage 185.
Bassit et al (Bassit RA, Pinheiro CH, Vitzel KF, Sproesser AJ, Silveira LR, Curi R. Effect of short-term creatine supplementation on markers of skeletal muscle damage after strenuous contractile activity. Eur J Appl Physiol. 2010;108:945–955. doi: 10.1007/s00421-009-1305-1. https://www.ncbi.nlm.nih.gov/pubmed/19956970()) observed a decrease in several markers of muscle damage (creatine kinase, lactate dehydrogenase, aldolase, glutamic oxaloacetic acid transaminase and glutamic pyruvic acid transaminase) in 4 athletes after an iron man competition who supplemented with 20 g/d plus 50 g maltodextrin during a 5 day period prior to the competition.
Cooke et al 186 observed positive effects of a prior (0.3 g/d kg body weight) loading and a post maintenance protocol (0.1 g/d kg body weight) to attenuate the loss of strength and muscle damage after an acute supramaximal (3 set x 10 rep with 120% 1RM) eccentric resistance training session in young males. The authors speculate that creatine ingestion prior to exercise may enhance calcium buffering capacity of the muscle and reduce calcium-activated proteases which in turn minimize sarcolemma and further influxes of calcium into the muscle. In addition creatine ingestion post exercise would enhance regenerative responses, favoring a more anabolic environment to avoid severe muscle damage and improve the recovery process. In addition, in vitro studies have demonstrated the antioxidant effects of creatine to remove superoxide anion radicals and peroxinitrite radicals 187. This antioxidant effect of creatine has been associated with the presence of Arginine in its molecule. Arginine is also a substrate for nitric oxide synthesis and can increase the production of nitric oxide which has higher vasodilatation properties, and acts as a free radical that modulates metabolism, contractibility and glucose uptake in skeletal muscle. Other amino acids contained in the creatine molecule such as glycine and methinine may be especially susceptible to free radical oxidation because of sulfhydryl groups 187. A more recent in vitro study showed that creatine exerts direct antioxidant activity via a scavenging mechanism in oxidatively injured cultured mammalian cells 188. In a recent in vivo study Rhaini et al 189 showed a positive effect of 7 days of creatine supplementation (4 x 5 g CM 20 g total) on 27 recreational resistance trained males to attenuate the oxidation of DNA and lipid peroxidation after a strenuous resistance training protocol.
Collectively the above investigations indicate that creatine supplementation can be an effective strategy to maintain total creatine pool during a rehabilitation period after injury as well as to attenuate muscle damage induced by a prolonged endurance training session. In addition, it seems that creatine can act as an effective antioxidant agent after more intense resistance training sessions 157.
Effects of creatine supplementation on glycogen stores
It is suggested 190, 191 that another mechanism for the effect of creatine could be enhanced muscle glycogen accumulation and GLUT4 expression, when creatine supplementation is combined with a glycogen depleting exercise. Whereas it has been observed 192 that creatine supplementation alone does not enhance muscle glycogen storage. Hickner et al 193 observed positive effects of creatine supplementation for enhancing initial and maintaining a higher level of muscle glycogen during 2 hours of cycling. In general, it is accepted that glycogen depleting exercises, such as high intensity or long duration exercise should combine high carbohydrate diets with creatine supplementation to achieve heightened muscle glycogen stores 185.
Effects of creatine supplementation on predominantly aerobic exercise
Although creatine supplementation has been shown to be more effective on predominantly anaerobic intermittent exercise, there is some evidence of its positive effects on endurance activities. Branch 194 highlights that endurance activities lasting more than 150 seconds rely on oxidative phosphorylation as primary energy system supplier. From this meta analysis 194, it would appear that the ergogenic potential for creatine supplementation on predominantly aerobic endurance exercise diminishes as the duration of the activity increases over 150s. However it is suggested that creatine supplementation may cause a change in substrate utilization during aerobic activity possibly leading to an increase in steady state endurance performance.
Chwalbinska-Monteta 195 observed a significant decrease in blood lactate accumulation when exercising at lower intensities as well as an increase in lactate threshold in elite male endurance rowers after consuming a short loading (5 days 20 g/d) creatine monohydrate protocol. However, the effects of creatine supplementation on endurance performance have been questioned by some studies. Graef et al 196 examined the effects of four weeks of creatine citrate supplementation and high-intensity interval training on cardio respiratory fitness. A greater increase of the ventilatory threshold was observed in the creatine group respect to placebo; however, oxygen consumption showed no significant differences between the groups. The total work presented no interaction and no main effect for time for any of the groups. Thompson et al 197 reported no effects of a 6 week 2 g creatine monohydrate/day in aerobic and anaerobic endurance performance in female swimmers.
Documented effects of creatine supplementation on physical performance
The majority of studies focusing on creatine supplementation report an increase in the body’s’ creatine pool 193, 190, 178. There is a positive relationship between muscle creatine uptake and exercise performance 178. Volek et al. 198 observed a significant increase in strength performance after 12 weeks creatine supplementation with a concurrent periodized heavy resistance training protocol. The creatine supplementation protocol consisted of a weeklong loading period of 25 g/day followed by a 5 gram maintenance dose for the remainder of the training. These positive effects were attributed to an increased total creatine pool resulting in more rapid adenosine triphosphate (ATP) regeneration between resistance training sets allowing athletes to maintain a higher training intensity and improve the quality of the workouts along the entire training period.
It is regularly reported that creatine supplementation, when combined with heavy resistance training leads to enhanced physical performance, fat free mass, and muscle morphology 199, 200, 201. A 2003 meta analysis 174 showed individuals ingesting creatine, combined with resistance training, obtain on average +8% and +14% more performance on maximum (1RM) or endurance strength (maximal repetitions at a given percent of 1RM) respectively than the placebo groups. However, contradicting studies have reported no effects of creatine supplementation on strength performance. Jakobi et al 202 found no effects of a short term creatine loading protocol upon isometric elbow flexion force, muscle activation, and recovery process. However, this study did not clearly state if creatine supplementation was administered concurrent with resistance training. Bemben et al 203 have shown no additional benefits of creatine alone or combined with whey protein for improving strength and muscle mass after a progressive 14 weeks (3 days per week) resistance training program in older men. These conflicting results can be explained by the possibility that the supplemented groups were formed by a greater amount of non-responders or even because creatine supplementation was administered on the training days only (3 times a week). This strategy has not been adequately tested as effective in middle aged and older men for maintaining post loading elevated creatine stores 171.
A quantitative, comprehensive scientific summary and view of knowledge up to 2007 on the effects of creatine supplementation in athletes and active people was published in a 100 citation review position paper by the International Society of Sports Nutrition 171. More recent literature has provided greater insight into the anabolic/performance enhancing mechanisms of creatine supplementation 193, 204 suggesting that these effects may be due to satellite cell proliferation, myogenic transcription factors and insulin-like growth factor-1 signalling 190. Saremi et al 205 reported a change in myogenic transcription factors when creatine supplementation and resistance training are combined in young healthy males. It was found that serum levels of myostatin, a muscle growth inhibitor, were decreased in the creatine group.
Collectively, in spite of a few controversial results, it seems that creatine supplementation combined with resistance training would amplify performance enhancement on maximum and endurance strength as well muscle hypertrophy 157.
Effects of creatine supplementation on predominantly anaerobic exercise
Creatine has demonstrated neuromuscular performance enhancing properties on short duration, predominantly anaerobic, intermittent exercises. Bazzucch et al 206 observed enhanced neuromuscular function of the elbow flexors in both electrically induced and voluntary contractions but not on endurance performance after 4 loading doses of 5 g creatine plus 15 g maltodextrin for 5/d in young, moderately trained men. Creatine supplementation may facilitate the reuptake of Ca2+ into the sacroplasmic reticulum by the action of the Ca2+ adenosine triphosphatase pump, which could enable force to be produced more rapidly through the faster detachment of the actomyosin bridges.
A previous meta-analysis 194 reported an overall creatine supplementation effect size (ES) of 0.24 ± 0.02 for activities lasting ≤30 s. (primarily using the ATP- phosphocreatine energy system). For this short high-intensity exercise, creatine supplementation resulted in a 7.5 ± 0.7% increase from base line which was greater than the 4.3 ± 0.6% improvement observed for placebo groups. When looking at the individual selected measures for anaerobic performance the greatest effect of creatine supplementation was observed on the number of repetitions which showed an ES of 0.64 ± 0.18. Furthermore, an increase from base line of 45.4 ± 7.2% compared to 22.9 ± 7.3% for the placebo group was observed. The second greatest ES was on the weight lifted at 0.51 ± 0.16 with an increase from base line of 13.4 ± 2.7% for the placebo group and 24.7 ± 3.9% for the creatine group. Other measures improved by creatine with a mean ES greater than 0 were for the amount of work accomplished, weight lifted, time, force production, cycle ergometer revolutions/min and power. The possible effect of creatine supplementation on multiple high intensity short duration bouts (<30 s) have shown an ES not statistically significant from 0. This would indicate that creatine supplementation might be useful to attenuate fatigue symptoms over multiple bouts of high-intensity, short duration exercise. The ES of creatine on anaerobic endurance exercise (>30 – 150s), primarily using the anaerobic glycolysis energy system, was 0.19 ± 0.05 with an improvement from baseline of 4.9 ± 1.5 % for creatine and -2.0 ± 0.6% for the placebo. The specific aspects of anaerobic endurance performance improved by creatine supplementation were work and power, both of which had a mean ES greater than 0. From the findings of this previous meta-analysis 194 it would appear that creatine supplementation has the most pronounced effect on short duration (<30s) high intensity intermittent exercises.
Effects of creatine supplementation on predominantly aerobic exercise
Although creatine supplementation has been shown to be more effective on predominantly anaerobic intermittent exercise, there is some evidence of its positive effects on endurance activities. Branch 194 highlights that endurance activities lasting more than 150 seconds rely on oxidative phosphorylation as primary energy system supplier. From this meta analysis 194, it would appear that the ergogenic potential for creatine supplementation on predominantly aerobic endurance exercise diminishes as the duration of the activity increases over 150s. However it is suggested that creatine supplementation may cause a change in substrate utilization during aerobic activity possibly leading to an increase in steady state endurance performance.
Chwalbinska-Monteta 195 observed a significant decrease in blood lactate accumulation when exercising at lower intensities as well as an increase in lactate threshold in elite male endurance rowers after consuming a short loading (5 days 20 g/d) CM protocol. However, the effects of creatine supplementation on endurance performance have been questioned by some studies. Graef et al 196 examined the effects of four weeks of creatine citrate supplementation and high-intensity interval training on cardio respiratory fitness. A greater increase of the ventilatory threshold was observed in the creatine group respect to placebo; however, oxygen consumption showed no significant differences between the groups. The total work presented no interaction and no main effect for time for any of the groups. Thompson et al 197 reported no effects of a 6 week 2 g CM/d in aerobic and anaerobic endurance performance in female swimmers. In addition, of the concern related to the dosage used in these studies, it could be possible that the potential benefits of creatine supplementation on endurance performance were more related to effects of anaerobic threshold localization.
Effects of creatine supplementation on range of motion
Sculthorpe et al 207 has shown that a 5 day (25g/d) loading protocol of creatine supplementation followed by a further 3 days of 5 g/d negatively influence both active ankle dorsiflexion and shoulder abduction and extension range of movement (ROM) in young men. There are two possible theories to explain these effects: 1) Creatine supplementation increases intracellular water content resulting in increased muscle stiffness and resistance to stretch; 2) Neural outflow from the muscle spindles is affected due to an increased volume of the muscle cell. The authors highlight that the active ROM measures were taken immediately after the loading phase and the reduced active ROM may not be seen after several weeks of maintenance phase 207. Hile et al 208 observed an increase in compartment pressure in the anterior compartment of the lower leg, which may also have been responsible for a reduced active ROM.
Creatine use in children and adolescents
Creatine supplementation in the under 18 population has not received a great deal of attention, especially in regards to sports/exercise performance. Despite this, creatine is being supplemented in young, <18 years old, athletes 209, 210. In a 2001 report 209 conducted on pupils from middle and high school (aged 10 – 18) in Westchester County (USA) 62 of the 1103 pupils surveyed were using creatine. The authors found this concerning for 2 main reasons: firstly, the safety of creatine supplementation is not established for this age group and is therefore not recommended. Secondly, it was speculated that taking creatine would lead on to more dangerous performance enhancing products such as anabolic steroids. It is important to point out that this potential escalation is speculation. Furthermore, a questionnaire was used to determine creatine use amongst this age group and does not necessarily reflect the truth.
A child’s ability to regenerate high energy phosphates during high intensity exercise is less than that of an adult. Due to this, creatine supplementation may benefit the rate and use of creatine phosphate and ATP rephosporylation. However, performance in short duration high-intensity exercise can be improved through training therefore supplementation may not be necessary 211.
Based on the limited data on performance and safety, some authors have not identified any conclusions and do not recommend its consumption in regards to creatine supplementation in children and adolescents 210, 211. Conversely, according to the view of the International Society of Sports Nutrition 171, younger athletes should consider a creatine supplement under certain conditions: puberty is past and he/she is involved in serious competitive training; the athlete is eating a well-balanced caloric adequate diet; he/she as well as the parents approve and understand the truth concerning the effects of creatine supplementation; supplement protocols are supervised by qualified professionals; recommended doses must not be exceeded; quality supplements are administered.
Within this framework, creatine supplementation in young, post puberty athletes can be considered a high quality type of “food” that can offer additional benefits to optimise training outcomes.
Safety and side effects of creatine supplementation
There have been a few reported renal health disorders associated with creatine supplementation 212, 213. These are isolated reports in which recommended dosages are not followed or there is a history of previous health complaints, such as renal disease or those taking nephrotoxic medication aggravated by creatine supplementation 213. Specific studies into creatine supplementation, renal function and/or safety conclude that although creatine does slightly raise creatinine levels there is no progressive effect to cause negative consequences to renal function and health in already healthy individuals when proper dosage recommendations are followed 214, 215, 216, 213. Urinary methylamine and formaldehyde have been shown to increase due to creatine supplementation of 20 g/d; this however did not bring the production outside of normal healthy range and did not impact on kidney function 217, 218. It has been advised that further research be carried out into the effects of creatine supplementation and health in the elderly and adolescent 212, 216. More recently, a randomized, double blind, 6 month resistance exercise and supplementation intervention 219 was performed on elderly men and women (age >65 years) in which subjects were assigned to either a supplement or placebo group. The supplement group was given 5 g creatine monohydrate, 2 g dextrose and 6 g conjugated linoleic acid/d, whilst the placebo group consumed 7 g dextrose and 6 g safflower oil/d. Creatine monohydrate administration showed significantly greater effects to improve muscular endurance, isokinetic knee extension strength, fat free mass and to reduce fat mass compared to placebo. Furthermore the supplement group had an increase in serum creatinine but not creatinine clearance suggesting no negative effect on renal function.
Cornelissen et al 220 analyzed the effects of 1 week loading protocol (3 X 5 g/d creatine monohydrate) followed by a 3 month maintenance period (5 g/d) on cardiac patients involved in an endurance and resistance training program. Although creatine monohydrate supplementation did not significantly enhance performance, markers of renal and liver function were within normal ranges indicating the safety of the applied creatine supplementation protocol.
A retrospective study 221, that examined the effects of long lasting (0.8 to 4 years) creatine monohydrate supplementation on health markers and prescribed training benefits, suggested that there is no negative health effects (including muscle cramp or injuries) caused by long term creatine monohydrate consumption. In addition, despite many anecdotal claims, it appears that creatine supplementation would have positive influences on muscle cramps and dehydration 222. Creatine was found to increase total body water possibly by decreasing the risk of dehydration, reducing sweat rate, lowering core body temperature and exercising heart rate. Furthermore, creatine supplementation does not increase symptoms nor negatively affect hydration or thermoregulation status of athletes exercising in the heat 223, 224. Additionally, CM ingestion has been shown to reduce the rate of perceived exertion when training in the heat 225.
It is prudent to note that creatine supplementation has been shown to reduce the body’s endogenous production of creatine, however levels return to normal after a brief period of time when supplementation ceases 226, 172. Despite this creatine supplementation has not been studied/supplemented with for a relatively long period. Due to this, long term effects are unknown, therefore safety cannot be guaranteed. Whilst the long term effects of creatine supplementation remain unclear, no definitive certainty of either a negative or a positive effect upon the body has been determined for many health professionals and national agencies 227, 217. For example the French Sanitary Agency has banned the buying of creatine due to the unproven allegation that a potential effect of creatine supplementation could be that of mutagenicity and carcinogenicity from the production of heterocyclic amines 217. Long term and epidemiological data should continue to be produced and collected to determine the safety of creatine in all healthy individuals under all conditions 217.
Commercially available forms of creatine supplements
There are several different available forms of creatine: creatine anhydrous which is creatine with the water molecule removed in order to increase the concentration of creatine to a greater amount than that found in creatine monohydrate. Creatine has been manufactured in salt form: creatine pyruvate, creatine citrate, creatine malate, creatine phosphate, magnesium creatine, creatine oroate, Kre Alkalyn (creatine with baking soda). Creatine can also be manufactured in an ester form. Creatine ethyl ester (hydrochloride) is an example of this, as is creatine gluconate which is creatine bound to glucose. Another form is creatine effervescent which is creatine citrate or creatine monohydrate with citric acid and bicarbonate. The citric acid and bicarbonate react to produce an effervescent effect. When mixed with water the creatine separates from its carrier leaving a neutrally charged creatine, allowing it to dissolve to a higher degree in water. Manufacturers claim that creatine effervescent has a longer and more stable life in solution. When di-creatine citrate effervescent was studied 228 for stability in solution it was found that the di-creatine citrate dissociates to citric acid and creatine in aqueous solutions which in turn forms creatine monohydrate and eventually crystallises out of the solution due to its low solubility. Some of the creatine may also convert to creatinine.
Jager et al 229 observed 1.17 and 1.29 greater peak plasma creatine concentration 1 hour after ingesting creatine pyruvate compared to isomolar amount of creatine monohydrate and creatine citrate respectively. However time to peak concentration, and velocity constants of absorption and elimination, was the same for all three forms of creatine. Although not measured in this study it is questionable that these small differences in plasma creatine concentrations would have any effect on the increase of muscle creatine uptake. Jäger et al 230 investigated the effects of 28-days of creatine pyruvate and citrate supplementation on endurance capacity and power measured during an intermittent handgrip (15 seconds effort per 45 seconds rest) exercise in healthy young athletes. The authors used a daily dose protocol with the intention to slowly saturate muscle creatine stores. Both forms of creatine showed slightly different effects on plasma creatine absorption and kinetics. The two creatine salts significantly increased mean power but only pyruvate forms showed significant effects for increasing force and attenuating fatigability during all intervals. These effects can be attributed to an enhanced contraction and relaxation velocity as well as a higher blood flow and muscle oxygen uptake. On the other hand, the power performance measured with the citrate forms decreases with time and improvements were not significant during the later intervals. In spite of these positive trends further research is required about the effects of these forms of creatine as there is little or no evidence for their safety and efficacy. Furthermore the regularity status of the novel forms of creatine vary from country to country and are often found to be unclear when compared to that of creatine monohydrate 231.
In summary, creatine salts have been show to be less stable than creatine monohydrate. However the addition of carbohydrates could increase their stability 231. The potential advantages of creatine salts over creatine monohydrate include enhanced aqueous solubility and bioavailability which would reduce their possible gastrointestinal adverse effects 232. The possibility for new additional formulation such as tablets or capsules is interesting for its therapeutic application due to its attributed better dissolution kinetics and oral absorption compared to creatine monohydrate 232. However more complete in vivo pharmaceutical analysis of creatine salts are required to fully elucidate their potential advantages/disadvantages over the currently available supplement formulations.
Creatine is a hydrophilic polar molecule that consists of a negatively charged carboxyl group and a positively charged functional group 233. The hydrophilic nature of creatine limits its bioavailability [65]. In an attempt to increase creatines bioavailability creatine has been esterified to reduce the hydrophilicity; this product is known as creatine ethyl ester. Manufacturers of creatine ethyl ester promote their product as being able to by-pass the creatine transporter due to improved sarcolemmal permeability toward creatine 234. Spillane et al 234 analyzed the effects of a 5 days loading protocol (0.30 g/kg lean mass) followed by a 42 days maintenance phase (0.075 g/kg lean mass) of CM or ethyl ester both combined with a resistance training program in 30 novice males with no previous resistance training experience. The results of this study 234 showed that ethyl ester was not as effective as creatine monohydrate to enhance serum and muscle creatine stores. Furthermore creatine ethyl ester offered no additional benefit for improving body composition, muscle mass, strength, and power. This research did not support the claims of the creatine ethyl ester manufacturers.
Polyethylene glycol is a non-toxic, water-soluble polymer that is capable of enhancing the absorption of creatine and various other substances 235. Polyethylene glycol can be bound with creatine monohydrate to form polyethylene glycosylated creatine. One study 236 found that 5 g/d for 28 days of polyethylene glycosylated creatine was capable of increasing 1RM bench press in 22 untrained young men but not for lower body strength or muscular power. Body weight also did not significantly change in the creatine group which may be of particular interest to athletes in weight categories that require upper body strength. Herda et al 237 analyzed the effects of 5 g of creatine monohydrate and two smaller doses of polyethylene glycosylated creatine (containing 1.25 g and 2.5 g of creatine) administered over 30 days on muscular strength, endurance, and power output in fifty-eight healthy men. Creatine monohydrate produced a significantly greater improvement in mean power and body weight meanwhile both creatine monohydrate and polyethylene glycosylated form showed a significantly greater improvement for strength when compared with control group. These strength increases were similar even though the dose of creatine in the polyethylene glycosylated creatine groups was up to 75% less than that of creatine monohydrate. These results seem to indicate that the addition of polyethylene glycol could increase the absorption efficiency of creatine but further research is needed before a definitive recommendation can be reached.
Creatine in combination with other supplements
Although creatine can be bought commercially as a standalone product it is often found in combination with other nutrients. A prime example is the combination of creatine with carbohydrate or protein and carbohydrate for augmenting creatine muscle retention 171 mediated through an insulin response from the pancreas 238. Steenge et al 239 found that body creatine retention of 5 g CM was increased by 25% with the addition of 50 g of protein and 47 g of carbohydrate or 96 g carbohydrate when compared to a placebo treatment of 5 g carbohydrate. The addition of 10g of creatine to 75 g of dextrose, 2 g of taurine, vitamins and minerals, induced a change in cellular osmolarity which in addition to the expected increase in body mass, seems to produce an up regulation of large scale gene expression (mRNA content of genes and protein content of kinases involved in osmosensing and signal transduction, cytoskeleton remodelling, protein and glycogen synthesis regulation, satellite cell proliferation and differentiation, DNA replication and repair, RNA transcription control, and cell survival) 204. Similar findings have also been reported for creatine monohydrate supplementation alone when combined with resistance training 240.
A commercially available pre-workout formula comprised of 2.05 g of caffeine, taurine and glucuronolactone, 7.9 g of L-leucine, L-valine, L-arginine and L-glutamine, 5 g of di-creatine citrate and 2.5 g of β-alanine mixed with 500 ml of water taken 10 minutes prior to exercise has been shown to enhance time to exhaustion during moderate intensity endurance exercise and to increase feelings of focus, energy and reduce subjective feelings of fatigue before and during endurance exercise due to a synergistic effect of the before mentioned ingredients 241. The role of creatine in this formulation is to provide a neuroprotective function by enhancing the energy metabolism in the brain tissue, promoting antioxidant activities, improving cerebral vasculation and protecting the brain from hyperosmotic shock by acting as a brain cell osmolyte. Creatine can provide other neuroprotective benefits through stabilisation of mitochondrial membranes, stimulation of glutamate uptake into synaptic vesicles and balance of intracellular calcium homeostasis 241.
Summary
The above review indicates that creatine supplementation has positive effects on:
- Amplifying the effects of resistance training for enhancing strength and muscle mass.
- Improving the quality and benefits of high intensity intermittent speed training.
- Improving aerobic endurance performance in trials lasting more than 150s.
- Seems to produce positive effects on strength, power, fat free mass, daily living performance and neurological function in young and older people.
- Regarding predominantly aerobic endurance performance, the increased bodies’ creatine stores, seems to amplify favorable physiological adaptations such as: increased plasma volume, glycogen storage, improvements of ventilatory threshold and a possible reduction of oxygen consumption in sub maximal exercise.
A typical creatine supplementation protocol of either a loading phase of 20 to 25 g CM/d or 0.3 g CM/kg/d split into 4 to 5 daily intakes of 5 g each have been recommended to quickly saturate creatine stores in the skeletal muscle. However a more moderate protocol where several smaller doses of creatine are ingested along the day (20 intakes of 1 g every 30 min) could be a better approach to get a maximal saturation of the intramuscular creatine store. In order to keep the maximal saturation of body creatine, the loading phase must be followed by a maintenance period of 3-5 g CM/d or 0.03 g CM/kg/d. These strategies appear to be the most efficient way of saturating the muscles and benefitting from CM supplementation. However more recent research has shown CM supplementation at doses of 0.1 g/kg body weight combined with resistance training improves training adaptations at a cellular and sub-cellular level. Creatine retention by the body from supplementation appears to be promoted by about 25% from the simultaneous ingestion of carbohydrate and/or protein mediated through an increase in insulin secretion. This combination would produce a faster saturation rate but has not been shown to have a greater effect on performance.
The available evidence indicates that creatine consumption is safe. This perception of safety cannot be guaranteed especially that of the long term safety of creatine supplementation and the various forms of creatine which are administered to different populations (athletes, sedentary, patient, active, young or elderly) throughout the world.
Branched-Chain Amino Acid Supplement
It is well-established that ingestion of essential amino acids following resistance exercise stimulates an increased response of muscle protein synthesis (MPS) in humans 242, 243, 244. Indeed, the stimulation of muscle protein synthesis in humans can be achieved by supplying essential amino acids only (i.e., the non-EAAs necessary for MPS may be supplied by endogenous sources) 245. More recent evidence from studies in rodents and cell culture models suggest that the stimulation of muscle protein synthesis by essential amino acids may be mediated by a few amino acids rather than a combination of all essential amino acids 246. The branched-chain amino acid (BCAA), leucine, has been shown to play a unique role in stimulating MPS 246. Leucine serves as substrate for the synthesis of new muscle proteins and as a signal to initiate the rate-limiting translation initiation step of muscle protein synthesis 247. Accordingly, the response of MPS to leucine provision has been extensively investigated over the past two decades, both in cell culture studies and in vivo rodent and human 248 studies.
The stimulation of muscle protein synthesis is accompanied by an increased activation of intracellular signaling proteins that regulate the translational activity of muscle protein synthesis 249. In particular, the mechanistic/mammalian target of rapamycin complex-1 (mTORC1) signaling, often assessed as the phosphorylation status of the ribosomal S6 protein kinase (S6K1), is stimulated by ingestion of EAA following resistance exercise 250. There is current debate over whether leucine alone 251, or the BCAAs combined 250, provide the most important component of an exogenous EAA source for stimulating the mTORC1-S6K1 signaling pathway. Whereas, the inclusion of leucine is necessary for the maximal activation of mTORC1 signaling 252, recent results from the same research group show that mTORC1 signaling is enhanced with the addition of the other two BCAAs, valine and isoleucine 250. Moreover, there often is a disconnect between the response of mTORC1 signaling and muscle protein synthesis 253, 254. Thus, the response of MPS to BCAA ingestion is still uncertain.
Branched chain amino acids
Branched chain amino acids (BCAA’s) make up 14-18% of amino acids in skeletal muscle proteins and are quite possibly the most widely used supplements among natural bodybuilders 255. Of the BCAA’s, leucine is of particular interest because it has been shown to stimulate protein synthesis to an equal extent as a mixture of all amino acids 256. However, ingestion of leucine alone can lead to depletion of plasma valine and isoleucine; therefore, all three amino acids need to be consumed to prevent plasma depletion of any one of the BCAA’s 257. Recently, the safe upper limit of leucine was set at 550 mg/kg bodyweight/day in adult men; however, future studies are needed to determine the safe upper limit for both other populations and a mixture of all 3 BCAA’s 258.
Numerous acute studies in animals and humans have shown that consumption of either essential amino acids, BCAA’s, or leucine either at rest or following exercise increases skeletal muscle protein synthesis, decreases muscle protein degradation, or both 259, 260; however, there are few long-term studies of BCAA supplementation in resistance-trained athletes. Stoppani et al. 261 supplemented trained subjects with either 14 g BCAAs, whey protein, or a carbohydrate placebo for eight weeks during a periodized strength training routine. After training the BCAA group had a 4 kg increase in lean mass, 2% decrease in body fat percentage, and 6 kg increase in bench press 10 repetition maximum. All changes were significant compared to the other groups. However, it should be noted that this data is only available as an abstract and has yet to undergo the rigors of peer-review.
The use of BCAA’s between meals may also be beneficial to keep protein synthesis elevated. Recent data from animal models suggest that consumption of BCAA’s between meals can overcome the refractory response in protein synthesis that occurs when plasma amino acids are elevated, yet protein synthesis is reduced 262. However, long-term human studies examining the effects of a diet in which BCAA’s are consumed between meals on lean mass and strength have not been done to date. It should also be noted that BCAA metabolism in humans and rodents differ and the results from rodent studies with BCAA’s may not translate in human models 263. Therefore, long-term studies are needed in humans to determine the effectiveness of this practice.
Based on the current evidence, it is clear BCAA’s stimulate protein synthesis acutely and one study 261 has indicated that BCAA’s may be able to increase lean mass and strength when added to a strength training routine; however, additional long-term studies are needed to determine the effects of BCAA’s on lean mass and strength in trained athletes. In addition, studies are needed on the effectiveness of BCAA supplementation in individuals following a vegetarian diet in which consumption of high-quality proteins are low as this may be population that may benefit from BCAA consumption. Furthermore, the effects of BCAA ingestion between meals needs to be further investigated in a long-term human study.
Arginine
“NO supplements” containing arginine are consumed by bodybuilders pre-workout in an attempt to increase blood flow to the muscle during exercise, increase protein synthesis, and improve exercise performance. However, there is little scientific evidence to back these claims. Fahs et al. 264 supplemented healthy young men with 7 g arginine or a placebo prior to exercise and observed no significant change in blood flow following exercise. Additionally, Tang et al. 265 supplemented either 10 g arginine or a placebo prior to exercise and found no significant increase in blood flow or protein synthesis following exercise. Moreover, arginine is a non essential amino acid and prior work has established that essential amino acids alone stimulate protein synthesis 266. Based on these findings, it appears that arginine does not significantly increase blood flow or enhance protein synthesis following exercise.
The effects of arginine supplementation on performance are controversial. Approximately one-half of acute and chronic studies on arginine and exercise performance have found significant benefits with arginine supplementation, while the other one-half has found no significant benefits 267. Moreover, Greer et al. 268 found that arginine supplementation significantly reduced muscular endurance by 2–4 repetitions on chin up and push up endurance tests. Based on these results, the authors of a recent review concluded that arginine supplementation had little impact on exercise performance in healthy individuals 269. Although the effects of arginine on blood flow, protein synthesis, and exercise performance require further investigation, dosages commonly consumed by athletes are well below the observed safe level of 20 g/d and do not appear to be harmful 270.
Citrulline malate
Citrulline malate (CitM) has recently become a popular supplement among bodybuilders; however, there has been little scientific research in healthy humans with this compound. CitM is hypothesized to improve performance through three mechanisms: 1) citrulline is important part of the urea cycle and may participate in ammonia clearance, 2) malate is a tricarboxylic acid cycle intermediate that may reduce lactic acid accumulation, and 3) citrulline can be converted to arginine; however, as discussed previously, arginine does not appear to have an ergogenic effect in young healthy athletes so it is unlikely CitM exerts an perfromance enhancing effect through this mechanism 271.
Supplementation with CitM for 15 days has been shown to increase ATP production by 34% during exercise, increase the rate of phosphocreatine recovery after exercise by 20%, and reduce perceptions of fatigue [184]. Moreover, ingestion of 8 g CitM prior to a chest workout significantly increased repetitions performed by approximately 53% and decreased soreness by 40% at 24 and 48 hours post-workout 271. Furthermore, Stoppani et al. 261 in an abstract reported a 4 kg increase in lean mass, 2 kg decrease in body fat percentage, and a 6 kg increase in 10 repetition maximum bench press after consumption of a drink containing 14 g BCAA, glutamine, and CitM during workouts for eight weeks; although, it is not clear to what degree CitM contributed to the outcomes observed. However, not all studies have supported ergogenic effects of CitM. Sureda et al. 272 found no significant difference in race time when either 6 g CitM or a placebo were consumed prior to a 137 km cycling stage. Hickner et al. 273 found that treadmill time to exhaustion was significantly impaired, with the time taken to reach exhaustion occurring on average seven seconds earlier following CitM consumption.
Additionally, the long-term safety of CitM is unknown. Therefore, based on the current literature a decision on the efficacy of CitM cannot be made. Future studies are needed to conclusively determine if CitM is ergogenic and to determine its long term safety.
Glutamine
Glutamine is the most abundant non-essential amino acid in muscle and is commonly consumed as a nutritional supplement. Glutamine supplementation in quantities below 14 g/d appear to be safe in healthy adults 270; however, at present there is little scientific evidence to support the use of glutamine in healthy athletes 274. Acutely, glutamine supplementation has not been shown to significantly improve exercise performance 275, 276, improve buffering capacity 276, help to maintain immune function or reduce muscle soreness after exercise 274. Long-term supplementation studies including glutamine in cocktails along with creatine monohydrate, whey protein, BCAA’s, and/or CitM have shown 1.5 – 2 kg increases in lean mass and 6 kg increase in 10RM bench press strength 261, 277. However, the role of glutamine in these changes is unclear. Only one study 278 has investigated the effects of glutamine supplementation alone in conjunction with a six week strength training program. No significant differences in muscle size, strength, or muscle protein degradation were observed between groups. Although the previous studies do not support the use of glutamine in bodybuilders during contest preparation, it should be noted that glutamine may be beneficial for gastrointestinal health and peptide uptake in stressed populations 279; therefore, it may be beneficial in dieting bodybuilders who represent a stressed population. As a whole, the results of previous studies do not support use of glutamine as an ergogenic supplement; however, future studies are needed to determine the role of glutamine on gastrointestinal health and peptide transport in dieting bodybuilders.
Whey protein
Milk protein is mostly composed of whey protein 20% and casein 80% 280. During cheese manufacturing, whey protein is generated as a by-product of casein precipitation. Whey protein is the most popular protein supplement sold in powder format. It contains valuable food ingredients because of its nutritional value and functional bioactivity. Whey protein contains β-lactoglobulin, α-lactalbumin, immunoglobulins, bovine serum albumin, lactoferrin, lactoperoxidase, phospholipoprotein, bioactive factors, and enzymes in order of abundance 281. The biological components of whey protein and its isolates have been reported to benefit antioxidation 282 and regulation of lipid metabolism 283 and have antifatigue 284 and antidiabetic properties 285.
Whey protein isolates contain enriched essential amino acids, including branched chain amino acids, which the body needs for tissue synthesis, energy, and health. The high leucine content (50%–75% more than other protein sources), one of the branched chain amino acids, in whey protein could explain its ability to stimulate muscle protein synthesis 286 and upregulate mammalian target of rapamycin signaling in high concentration. With whey protein supplementation, resistance exercises can result in muscle adaption and hypertrophy, regardless of the contraction mode (Whey protein hydrolysate augments tendon and muscle hypertrophy independent of resistance exercise contraction mode. Farup J, Rahbek SK, Vendelbo MH, Matzon A, Hindhede J, Bejder A, Ringgard S, Vissing K. Scand J Med Sci Sports. 2014 Oct; 24(5):788-98. https://www.ncbi.nlm.nih.gov/pubmed/23647357/()). Whey protein is marketed as a dietary supplement and as an aid for muscle development with resistance training. Because of its rapid rate of digestion, whey protein provides a rapid source of amino acids that can be taken up by the muscles to repair and rebuild muscular tissue. The use of whey protein to enhance aerobic exercises and swimming training has only been reported in terms of glycogen storage 287, antioxidation 288, and lipid metabolism. A combination of resistant exercise and WP benefitted the lipid profile, especially plasma triglycerides and cholesterol 289. Few reports have shown the beneficial synergistic effects of whey protein and long-term aerobic exercise training on biochemical profiles in specific tissues.
Resistance exercise, eccentric (muscle lengthening), isometric (non-lengthening) and concentric (shortening) contractions cause skeletal muscle damage and generate inflammatory markers (muscle proteins in blood) (The exercise-induced stress response of skeletal muscle, with specific emphasis on humans. Morton JP, Kayani AC, McArdle A, Drust B. Sports Med. 2009; 39(8):643-62. https://www.ncbi.nlm.nih.gov/pubmed/19769414/()). Anabolic interventions with protein hydrolysates and amino acid supplements have been evidenced to expedite the repair. Leucine-derived metabolite β-hydroxy-β-methylbutyrate ingestion has proved beneficial in recovery from the soreness. Resistance exercise (weight-lifting) elevates oxidation products in plasma, perturbs leukocyte redistribution and leukocyte functionality. Whey protein isolate nanoparticles were prepared using ethanol desolvation and their capacity to incorporate ZnCl2 was analysed. The amount of zinc incorporated in the particle suspensions was within the range of daily zinc requirements for healthy adults. Also, the nanoparticles remained stable after 30 days of storage at 22 °C. Cell surface glucose transporter 4 (GLUT 4) is the major glucose transporter isoform expressed in skeletal muscle that determines the rate of muscle glucose transport in the cell membrane, in response to insulin and muscle contraction. Whey protein hydrolysate was evaluated for its ability to translocate GLUT 4 and accumulate them in the membrane thereby augmenting glucose trapping by skeletal muscle. The amino acid l-isoleucine and the peptide l-leucyl-l-isoleucine in the hydrolysate contributed the most 290. The effect of whey supplementation in comparison to casein diet, on the recovery of muscle functional properties such as contractility, extensibility, elasticity and excitability was investigated in rats. The whey protein diet promoted a faster recovery from injury sustained due to isometric as well as concentric exercise in comparison to the casein diet 291. The effect of the supplementation of a beverage with varying doses of leucine or a mixture of branched chain amino acids on myofibrillar protein synthesis after resistance exercise was assessed. Results showed that low-protein (6.25 g) beverage can be as effective as a high-protein dose (25 g) at stimulating myofibrillar protein synthesis rates when supplemented with a high (5 g) leucine content. As leucine comprises 10 % of the total whey amino acid, the latter appears important for augmenting muscle hypertrophy. Health parameters, performance and body composition effects produced by 12 week intake of hydrolysed whey protein were compared in players. Intervention with the hydrolysed whey protein resulted in significant reduction in the muscle damage markers (creatine kinase and lactate dehydrogenase). Muscle mass growth by daily consumption of whey protein was compared with that of soy protein, using a randomized study on subjects undergoing resistance exercise 292. Lean body mass gains were significantly high in whey protein than soy protein group and the remarkable response was correlated with the elevated levels of leucine and faster absorption.
Concerning lean body mass, many studies reported that protein synthesis could be upregulated by the branched chain amino acids of whey protein, especially leucine 293. The combination of daily supplementation with whey protein and resistance exercise training was effective in promoting muscle hypertrophy 294.
Whey protein consumption at intake levels up to 3 g/kg per day had a no-observed-adverse-effect level 295 and the hydrolysate of whey protein at 2 g/kg as a food additive resulted in no adverse effects or mortality 296. In this study, the whey protein dose was 4.1 g/kg, which is equivalent to 20 g of whey protein per 60 kg body weight for humans did not reveal any adverse effects 297.
Based on their concentration and attributes, whey proteins are marketed in various forms such as whey protein concentrate (has fat and lactose along with proteins (29–89 %)), whey protein isolate (90 % protein) and whey protein hydrolysate (partially digested for ease of metabolism and hypo-allergenicity) 298. A broad range of functionality has been assigned to whey protein and its derivatives, such as reduction of oxidative stress, promotion of muscle growth and lean body mass, appetite suppression, hypoglycemia, cardiovascular risk mitigation, phenylketonuria management and protection from ultraviolet (UV) radiation 299. Further, its role in food processing such as emulsifier, texturizer, fat-replacer, encapsulating agent, delivery vehicle and antimicrobial film are being recognized 300.
What are Anabolic Steroids ?
Anabolic steroids are synthetic substances related to the male sex hormone testosterone. The proper term for these compounds is anabolic-androgenic steroids. “Anabolic” refers to muscle building, and “androgenic” refers to increased male sex characteristics. Some common names for anabolic steroids are Gear, Juice, Roids, and Stackers. They promote the growth of skeletal muscle (anabolic effects) and the development of male sexual characteristics (androgenic effects) in both males and females 301. The term “anabolic steroids” will be used throughout this report because of its familiarity, although the proper term for these compounds is “anabolic-androgenic steroids.”
Anabolic steroids were developed in the late 1930s primarily to treat hypogonadism, a condition in which the testes do not produce sufficient testosterone for normal growth, development, and sexual functioning. The primary medical uses of these compounds are to treat delayed puberty, some types of impotence, and wasting of the body caused by HIV infection or other diseases.
During the 1930s, scientists discovered that anabolic steroids could facilitate the growth of skeletal muscle in laboratory animals, which led to abuse of the compounds first by bodybuilders and weightlifters and then by athletes in other sports. Steroid abuse has become so widespread in athletics that it can affect the outcome of sports contests.
Illicit steroids are often sold at gyms, competitions, and through mail order operations after being smuggled into this country. Most illegal steroids in the United States are smuggled from countries that do not require a prescription for the purchase of steroids. Steroids are also illegally diverted from U.S. pharmacies or synthesized in clandestine laboratories.
What are steroidal supplements ?
In the United States, supplements such as tetrahydrogestrinone (THG) and androstenedione (street name “Andro”) previously could be purchased legally without a prescription through many commercial sources, including health food stores 302. Steroidal supplements can be converted into testosterone or a similar compound in the body. Less is known about the side effects of steroidal supplements, but if large quantities of these compounds substantially increase testosterone levels in the body, then they also are likely to produce the same side effects as anabolic steroids themselves. The purchase of these supplements, with the notable exception of dehydroepiandrosterone (DHEA), became illegal after the passage in 2004 of amendments to the Controlled Substances Act.
Commonly Abused Steroids
Oral Steroids
- Anadrol (oxymetholone)
- Oxandrin (oxandrolone)
- Dianabol (methandrostenolone)
- Winstrol (stanozolol)
Injectable Steroids
- Deca-Durabolin (nandrolone decanoate)
- Durabolin (nandrolone phenpropionate)
- Depo-Testosterone (testosterone cypionate)
- Equipoise (boldenone undecylenate)
- Tetrahydrogestrinone (THG).
How are anabolic steroids abused ?
Some anabolic steroids are taken orally, others are injected intramuscularly and still others are provided in gels or creams that are applied to the skin 303. Doses taken by abusers can be 10 to 100 times higher than the doses used for medical conditions 303.
Cycling, stacking, and pyramiding
Steroids are often abused in patterns called “cycling,” which involve taking multiple doses of steroids over a specific period of time, stopping for a period, and starting again 303. Users also frequently combine several different types of steroids in a process known as “stacking.” Steroid abusers typically “stack” the drugs, meaning that they take two or more different anabolic steroids, mixing oral and/or injectable types, and sometimes even including compounds that are designed for veterinary use 303. Abusers think that the different steroids interact to produce an effect on muscle size that is greater than the effects of each drug individually, a theory that has not been tested scientifically 303.
Another mode of steroid abuse is referred to as “pyramiding.” This is a process in which users slowly escalate steroid abuse (increasing the number of steroids or the dose and frequency of one or more steroids used at one time), reaching a peak amount at mid-cycle and gradually tapering the dose toward the end of the cycle 303. Often, steroid abusers pyramid their doses in cycles of 6 to 12 weeks. At the beginning of a cycle, the person starts with low doses of the drugs being stacked and then slowly increases the doses. In the second half of the cycle, the doses are slowly decreased to zero. This is sometimes followed by a second cycle in which the person continues to train but without drugs. Abusers believe that pyramiding allows the body time to adjust to the high doses, and the drug-free cycle allows the body’s hormonal system time to recuperate. As with stacking, the perceived benefits of pyramiding and cycling have not been substantiated scientifically 303.
Why do people abuse anabolic steroids ?
One of the main reasons people give for abusing steroids is to improve their athletic performance 304. Among athletes, steroid abuse has been estimated to be less that 6 percent according to surveys, but anecdotal information suggests more widespread abuse. Although testing procedures are now in place to deter steroid abuse among professional and Olympic athletes, new designer drugs constantly become available that can escape detection and put athletes willing to cheat one step ahead of testing efforts. This dynamic, however, may be about to shift if the saving of urine and blood samples for retesting at a future date becomes the standard. The high probability of eventual detection of the newer designer steroids, once the technology becomes available, plus the fear of retroactive sanctions, should give athletes pause.
Another reason people give for taking steroids is to increase their muscle size or to reduce their body fat 304. This group includes people suffering from the behavioral syndrome called muscle dysmorphia, which causes them to have a distorted image of their bodies. Men with muscle dysmorphia think that they look small and weak, even if they are large and muscular. Similarly, women with this condition think that they look fat and flabby, even though they are actually lean and muscular.
Some people who abuse steroids to boost muscle size have experienced physical or sexual abuse 304. In one series of interviews with male weightlifters, 25 percent who abused steroids reported memories of childhood physical or sexual abuse. Similarly, female weightlifters who had been raped were found to be twice as likely to report use of anabolic steroids or another purported musclebuilding drug, compared with those who had not been raped. Moreover, almost all of those who had been raped reported that they markedly increased their bodybuilding activities after the attack. They believed that being bigger and stronger would discourage further attacks because men would find them either intimidating or unattractive.
Finally, some adolescents abuse steroids as part of a pattern of high-risk behaviors 304. These adolescents also take risks such as drinking and driving, carrying a gun, driving a motorcycle without a helmet, and abusing other illicit drugs. Conditions such as muscle dysmorphia, a history of physical or sexual abuse, or a history of engaging in high-risk behaviors have all been associated with an increased risk of initiating or continuing steroid abuse.
Are anabolic steroids addictive ?
An undetermined percentage of steroid abusers may become addicted to the drugs, as evidenced by their continued abuse despite physical problems and negative effects on social relations. Also, steroid abusers typically spend large amounts of time and money obtaining the drugs, which is another indication that they may be addicted. Individuals who abuse steroids can experience withdrawal symptoms when they stop taking steroids, such as mood swings, fatigue, restlessness, loss of appetite, insomnia, reduced sex drive, and steroid cravings. The most dangerous of the withdrawal symptoms is depression, because it sometimes leads to suicide attempts. If left untreated, some depressive symptoms associated with anabolic steroid withdrawal have been known to persist for a year or more after the abuser stops taking the drugs.
People who abuse steroids may experience withdrawal symptoms when they stop use, including:
- mood swings
- fatigue
- restlessness
- loss of appetite
- sleep problems
- decreased sex drive
- steroid cravings.
One of the more serious withdrawal symptoms is depression, which can sometimes lead to suicide attempts.
What are the health consequences of steroid abuse ?
Anabolic steroid abuse has been associated with a wide range of adverse side effects ranging from some that are physically unattractive, such as acne and breast development in men, to others that are life threatening, such as heart attacks and liver cancer. Most are reversible if the abuser stops taking the drugs, but some are permanent, such as voice deepening in females.
Most data on the long-term effects of anabolic steroids in humans come from case reports rather than formal epidemiological studies. From the case reports, the incidence of lifethreatening effects appears to be low, but serious adverse effects may be underrecognized or underreported, especially since they may occur many years later. Data from animal studies seem to support this possibility. One study found that exposing male mice for one-fifth of their lifespan to steroid doses comparable to those taken by human athletes caused a high frequency of early deaths.
Anabolic steroids affect the brain
Anabolic steroids work differently from other drugs of abuse; they do not have the same short-term effects on the brain. The most important difference is that steroids do not trigger rapid increases in the brain chemical dopamine, which causes the “high” that drives people to abuse other substances. However, long-term steroid abuse can act on some of the same brain pathways and chemicals—including dopamine, serotonin, and opioid systems—that are affected by other drugs. This may result in a significant effect on mood and behavior.
Short-Term Effects
Abuse of anabolic steroids may lead to mental problems, such as:
- paranoid (extreme, unreasonable) jealousy
- extreme irritability
- delusions—false beliefs or ideas
- impaired judgment
Extreme mood swings can also occur, including “roid rage”—angry feelings and behavior that may lead to violence.
Aside from mental problems, steroid use commonly causes severe acne. It also causes the body to swell, especially in the hands and feet.
Long-Term Effects
Anabolic steroid abuse may lead to serious, even permanent, health problems such as:
- kidney problems or failure
- liver damage
- enlarged heart, high blood pressure, and changes in blood cholesterol, all of which increase the risk of stroke and heart attack, even in young people
Several other effects are gender- and age-specific:
In men:
- shrinking testicles
- decreased sperm count
- baldness
- development of breasts
- increased risk for prostate cancer.
In women:
- growth of facial hair or excess body hair
- male-pattern baldness
- changes in or stop in the menstrual cycle
- enlarged clitoris
- deepened voice.
In teens:
- stunted growth (when high hormone levels from steroids signal to the body to stop bone growth too early)
- stunted height (if teens use steroids before their growth spurt).
Some of these physical changes, such as shrinking sex organs in men, can add to mental side effects such as mood disorders.
Hormonal system
Steroid abuse disrupts the normal production of hormones in the body, causing both reversible and irreversible changes. Changes that can be reversed include reduced sperm production and shrinking of the testicles (testicular atrophy). Irreversible changes include male-pattern baldness and breast development (gynecomastia) in men. In one study of male bodybuilders, more than half had testicular atrophy and/or gynecomastia.
In the female body, anabolic steroids cause masculinization. Breast size and body fat decrease, the skin becomes coarse, the clitoris enlarges, and the voice deepens. Women may experience excessive growth of body hair but lose scalp hair. With continued administration of steroids, some of these effects become irreversible.
Musculoskeletal system
Rising levels of testosterone and other sex hormones normally trigger the growth spurt that occurs during puberty and adolescence and provide the signals to stop growth as well. When a child or adolescent takes anabolic steroids, the resulting artificially high sex hormone levels can prematurely signal the bones to stop growing.
Cardiovascular system
Steroid abuse has been associated with cardiovascular diseases (CVD), including heart attacks and strokes, even in athletes younger than 30. Steroids contribute to the development of CVD, partly by changing the levels of lipoproteins that carry cholesterol in the blood. Steroids, particularly oral steroids, increase the level of low-density lipoprotein (LDL) and decrease the level of high-density lipoprotein (HDL). High LDL and low HDL levels increase the risk of atherosclerosis, a condition in which fatty substances are deposited inside arteries and disrupt blood flow. If blood is prevented from reaching the heart, the result can be a heart attack. If blood is prevented from reaching the brain, the result can be a stroke.
Steroids also increase the risk that blood clots will form in blood vessels, potentially disrupting blood flow and damaging the heart muscle so that it does not pump blood effectively.
Liver
Steroid abuse has been associated with liver tumors and a rare condition called peliosis hepatis, in which blood-filled cysts form in the liver. Both the tumors and the cysts can rupture, causing internal bleeding.
Skin
Steroid abuse can cause acne, cysts, and oily hair and skin.
Infections
Many abusers who inject anabolic steroids may use nonsterile injection techniques or share contaminated needles with other abusers. In addition, some steroid preparations are manufactured illegally under nonsterile conditions. These factors put abusers at risk for acquiring lifethreatening viral infections, such as HIV and hepatitis B and C. Abusers also can develop endocarditis, a bacterial infection that causes a potentially fatal inflammation of the inner lining of the heart. Bacterial infections also can cause pain and abscess formation at injection sites.
What effects do anabolic steroids have on behavior ?
Case reports and small studies indicate that anabolic steroids, when used in high doses, increase irritability and aggression. Some steroid abusers report that they have committed aggressive acts, such as physical fighting or armed robbery, theft, vandalism, or burglary. Abusers who have committed aggressive acts or property crimes generally report that they engage in these behaviors more often when they take steroids than when they are drug free. A recent study suggests that the mood and behavioral effects seen during anabolic-androgenic steroid abuse may result from secondary hormonal changes.
Scientists have attempted to test the association between anabolic steroids and aggression by administering high steroid doses or placebo for days or weeks to human volunteers and then asking the people to report on their behavioral symptoms. To date, four such studies have been conducted. In three, high steroid doses did produce greater feelings of irritability and aggression than did placebo, although the effects appear to be highly variable across individuals. In one study, the drugs did not have that effect. One possible explanation, according to the researchers, is that some but not all anabolic steroids increase irritability and aggression. Recent animal studies show an increase in aggression after steroid administration.
In a few controlled studies, aggression or adverse, overt behaviors resulting from the administration of anabolic steroid use have been reported by a minority of volunteers.
In summary, the extent to which steroid abuse contributes to violence and behavioral disorders is unknown. As with the health complications of steroid abuse, the prevalence of extreme cases of violence and behavioral disorders seems to be low, but it may be underreported or underrecognized.
Research also indicates that some users might turn to other drugs to alleviate some of the negative effects of anabolic steroids. For example, a study of 227 men admitted in 1999 to a private treatment center for addiction to heroin or other opioids found that 9.3 percent had abused anabolic steroids before trying any other illicit drug. Of these 9.3 percent, 86 percent first used opioids to counteract insomnia and irritability resulting from anabolic steroids.
What treatments are effective for anabolic steroid abuse ?
Few studies of treatments for anabolic steroid abuse have been conducted. Current knowledge is based largely on the experiences of a small number of physicians who have worked with patients undergoing steroid withdrawal. The physicians have found that supportive therapy is sufficient in some cases. Patients are educated about what they may experience during withdrawal and are evaluated for suicidal thoughts. If symptoms are severe or prolonged, medications or hospitalization may be needed.
Some medications that have been used for treating steroid withdrawal restore the hormonal system after its disruption by steroid abuse. Other medications target specific withdrawal symptoms—for example, antidepressants to treat depression and analgesics for headaches and muscle and joint pains.
Some patients require assistance beyond pharmacological treatment of withdrawal symptoms and are treated with behavioral therapies.
Summary
Abuse of anabolic steroids differs from the abuse of other illicit substances because the initial abuse of anabolic steroids is not driven by the immediate euphoria that accompanies most drugs of abuse, such as cocaine, heroin, and marijuana, but by the desire of abusers to change their appearance and performance, characteristics of great importance to adolescents. The effects of steroids can boost confidence and strength, leading abusers to overlook the potential serious and long-term damage that these substances can cause.
While anabolic steroids can enhance certain types of performance or appearance, they are dangerous drugs, and when used inappropriately they can cause a host of severe, long-lasting, and in some cases, irreversible negative health consequences. Anabolic steroids can lead to early heart attacks, strokes, liver tumors, kidney failure, and serious psychiatric problems. In addition, because steroids are often injected, users who share needles or use nonsterile techniques when they inject steroids are at risk for contracting dangerous infections, such as HIV/AIDS and hepatitis B and C.
Possible Health Consequences of Anabolic Steroid Abuse
Hormonal system
Men
- infertility
- breast development
- shrinking of the testicles
- male-pattern baldness
Women
- enlargement of the clitoris
- excessive growth of body hair
- male-pattern baldness
Musculoskeletal system
- short stature (if taken by adolescents)
- tendon rupture
Cardiovascular system
- increases in LDL (bad) cholesterol;
- decreases in HDL (good) cholesterol;
- high blood pressure
- heart attacks
- enlargement of the heart’s left ventricle
Liver
- cancer
- peliosis hepatis (blood-filled cysts form in the liver)
- tumors
- Both the tumors and the cysts can rupture, causing internal bleeding.
Skin
- severe acne and cysts
- oily scalp
- jaundice
- fluid retention
Infection
- HIV/AIDS
- hepatitis B and C.
Psychiatric effects
- rage, aggression
- mania
- delusions
- Gundersen K. Excitation-transcription coupling in skeletal muscle: the molecular pathways of exercise. Biological Reviews of the Cambridge Philosophical Society. 2011;86(3):564-600. doi:10.1111/j.1469-185X.2010.00161.x. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3170710/
- Bintliff S, Walker BE. Radioautographic study of skeletal muscle regeneration. American Journal of Anatomy. 1960;106:233–245.
- Nature of dividing nuclei in skeletal muscle of growing rats. Moss FP, Leblond CP. J Cell Biol. 1970 Feb; 44(2):459-62. https://www.ncbi.nlm.nih.gov/pubmed/5411085/
- Morphological changes during fiber type transitions in low-frequency-stimulated rat fast-twitch muscle. Delp MD, Pette D. Cell Tissue Res. 1994 Aug; 277(2):363-71. https://www.ncbi.nlm.nih.gov/pubmed/7521794/
- Fast to slow transformation of denervated and electrically stimulated rat muscle. Windisch A, Gundersen K, Szabolcs MJ, Gruber H, Lømo T. J Physiol. 1998 Jul 15; 510 ( Pt 2)():623-32. https://www.ncbi.nlm.nih.gov/pubmed/9706009/
- Ferraro E, Giammarioli AM, Chiandotto S, Spoletini I, Rosano G. Exercise-Induced Skeletal Muscle Remodeling and Metabolic Adaptation: Redox Signaling and Role of Autophagy. Antioxidants & Redox Signaling. 2014;21(1):154-176. doi:10.1089/ars.2013.5773. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4048572/
- Schiaffino S. Fibre types in skeletal muscle: a personal account. Acta Physiol (Oxf) 199: 451–463, 2010. https://www.ncbi.nlm.nih.gov/pubmed/20353491
- Fibre types in skeletal muscle: a personal account. Schiaffino S. Acta Physiol (Oxf). 2010 Aug; 199(4):451-63. https://www.ncbi.nlm.nih.gov/pubmed/20353491/
- Burniston JG, Hoffman EP. Proteomic responses of skeletal and cardiac muscle to exercise. Expert review of proteomics. 2011;8(3):361-377. doi:10.1586/epr.11.17. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4948674/
- Molecular diversity of myofibrillar proteins: gene regulation and functional significance. Schiaffino S, Reggiani C. Physiol Rev. 1996 Apr; 76(2):371-423. https://www.ncbi.nlm.nih.gov/pubmed/8618961/
- Barany M. ATPase activity of myosin correlated with speed of muscle shortening. J Gen Physiol 50Suppl: 197–218, 1967. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2225740/pdf/197.pdf
- Schiaffino S. and Reggiani C. Molecular diversity of myofibrillar proteins: gene regulation and functional significance. Physiol Rev 76: 371–423, 1996. https://www.ncbi.nlm.nih.gov/pubmed/8618961
- Evolutionary implications of three novel members of the human sarcomeric myosin heavy chain gene family. Desjardins PR, Burkman JM, Shrager JB, Allmond LA, Stedman HH. Mol Biol Evol. 2002 Apr; 19(4):375-93. https://www.ncbi.nlm.nih.gov/pubmed/11919279/
- Hybrid skeletal muscle fibres: a rare or common phenomenon ? Stephenson GM. Clin Exp Pharmacol Physiol. 2001 Aug; 28(8):692-702. https://www.ncbi.nlm.nih.gov/pubmed/11473538/
- Functional heterogeneity of mammalian single muscle fibres: do myosin isoforms tell the whole story? Bottinelli R. Pflugers Arch. 2001 Oct; 443(1):6-17. https://www.ncbi.nlm.nih.gov/pubmed/11692261/
- PGC-1alpha regulation by exercise training and its influences on muscle function and insulin sensitivity. Lira VA, Benton CR, Yan Z, Bonen A. Am J Physiol Endocrinol Metab. 2010 Aug; 299(2):E145-61. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2928513/
- Excitation-transcription coupling in skeletal muscle: the molecular pathways of exercise. Gundersen K. Biol Rev Camb Philos Soc. 2011 Aug; 86(3):564-600. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3170710/
- Transitions of muscle fiber phenotypic profiles. Pette D, Staron RS. Histochem Cell Biol. 2001 May; 115(5):359-72. https://www.ncbi.nlm.nih.gov/pubmed/11449884/
- SERCA pump isoforms: their role in calcium transport and disease. Periasamy M, Kalyanasundaram A. Muscle Nerve. 2007 Apr; 35(4):430-42. https://www.ncbi.nlm.nih.gov/pubmed/17286271/
- Electrical stimulation resembling normal motor-unit activity: effects on denervated fast and slow rat muscles. Eken T, Gundersen K. J Physiol. 1988 Aug; 402():651-69. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1191914/
- Henriksson J, Hickner J. Training-induced adaptions in skeletal muscle. In: Harries M, editor. Oxford textbook of sports medicine. Oxford: Oxford University Press; 1994.
- Saltin B, Gollnick PD. Skeletal muscle adaptability: significance for metabolism and performance. In: Peachey LD, Adrian RH, Geiger SR, editors. Handbook of Physiology. Vol. 10. Bethesda: American Physiological Society; 1983. pp. 555–631
- Muscle mechanics: adaptations with exercise-training. Fitts RH, Widrick JJ. Exerc Sport Sci Rev. 1996; 24():427-73. https://www.ncbi.nlm.nih.gov/pubmed/8744258/
- Aerobic capacity and skeletal muscle properties of normoxic and hypoxic rats in response to training. Abdelmalki A, Fimbel S, Mayet-Sornay MH, Sempore B, Favier R. Pflugers Arch. 1996 Mar; 431(5):671-9.
- Compatibility of high-intensity strength and endurance training on hormonal and skeletal muscle adaptations. Kraemer WJ, Patton JF, Gordon SE, Harman EA, Deschenes MR, Reynolds K, Newton RU, Triplett NT, Dziados JE. J Appl Physiol (1985). 1995 Mar; 78(3):976-89. https://www.ncbi.nlm.nih.gov/pubmed/7775344/
- Myosin heavy chain isoforms in single fibres from m. vastus lateralis of sprinters: influence of training. Andersen JL, Klitgaard H, Saltin B. Acta Physiol Scand. 1994 Jun; 151(2):135-42. https://www.ncbi.nlm.nih.gov/pubmed/7942047/
- Myosin isoforms, muscle fiber types, and transitions. Pette D, Staron RS. Microsc Res Tech. 2000 Sep 15; 50(6):500-9. https://www.ncbi.nlm.nih.gov/pubmed/10998639/
- Novel transitions in MHC isoforms: separate and combined effects of thyroid hormone and mechanical unloading. Caiozzo VJ, Baker MJ, Baldwin KM. J Appl Physiol (1985). 1998 Dec; 85(6):2237-48. https://www.ncbi.nlm.nih.gov/pubmed/9843548/
- Cellular and molecular mechanisms responsible for the action of testosterone on human skeletal muscle. A basis for illegal performance enhancement. Kadi F. Br J Pharmacol. 2008 Jun; 154(3):522-8. https://www.ncbi.nlm.nih.gov/pubmed/18414389/
- Fiber type composition of the vastus lateralis muscle of young men and women. Staron RS, Hagerman FC, Hikida RS, Murray TF, Hostler DP, Crill MT, Ragg KE, Toma K. J Histochem Cytochem. 2000 May; 48(5):623-9. https://www.ncbi.nlm.nih.gov/pubmed/10769046/
- “Fast” and “slow” muscle fibres in hindlimb muscles of adult rats regenerate from intrinsically different satellite cells. Kalhovde JM, Jerkovic R, Sefland I, Cordonnier C, Calabria E, Schiaffino S, Lømo T. J Physiol. 2005 Feb 1; 562(Pt 3):847-57. https://www.ncbi.nlm.nih.gov/pubmed/15564285/
- Muscle fiber pattern is independent of cell lineage in postnatal rodent development. Hughes SM, Blau HM. Cell. 1992 Feb 21; 68(4):659-71. https://www.ncbi.nlm.nih.gov/pubmed/1531450/
- Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member. McPherron AC, Lawler AM, Lee SJ. Nature. 1997 May 1; 387(6628):83-90. https://www.ncbi.nlm.nih.gov/pubmed/9139826/
- Double muscling in cattle due to mutations in the myostatin gene. McPherron AC, Lee SJ. Proc Natl Acad Sci U S A. 1997 Nov 11; 94(23):12457-61. https://www.ncbi.nlm.nih.gov/pubmed/9356471/
- Myostatin mutation associated with gross muscle hypertrophy in a child. Schuelke M, Wagner KR, Stolz LE, Hübner C, Riebel T, Kömen W, Braun T, Tobin JF, Lee SJ. N Engl J Med. 2004 Jun 24; 350(26):2682-8. https://www.ncbi.nlm.nih.gov/pubmed/15215484/
- Lack of myostatin results in excessive muscle growth but impaired force generation. Amthor H, Macharia R, Navarrete R, Schuelke M, Brown SC, Otto A, Voit T, Muntoni F, Vrbóva G, Partridge T, Zammit P, Bunger L, Patel K. Proc Natl Acad Sci U S A. 2007 Feb 6; 104(6):1835-40. https://www.ncbi.nlm.nih.gov/pubmed/17267614/
- Muscle hypertrophy induced by the Ski protein: cyto-architecture and ultrastructure. Bruusgaard JC, Brack AS, Hughes SM, Gundersen K. Acta Physiol Scand. 2005 Oct; 185(2):141-9. https://www.ncbi.nlm.nih.gov/pubmed/16168008/
- Short-term strength training and the expression of myostatin and IGF-I isoforms in rat muscle and tendon: differential effects of specific contraction types. Heinemeier KM, Olesen JL, Schjerling P, Haddad F, Langberg H, Baldwin KM, Kjaer M. J Appl Physiol (1985). 2007 Feb; 102(2):573-81. https://www.ncbi.nlm.nih.gov/pubmed/17038487/
- Myostatin and insulin-like growth factor-I and -II expression in the muscle of rats exposed to the microgravity environment of the NeuroLab space shuttle flight. Lalani R, Bhasin S, Byhower F, Tarnuzzer R, Grant M, Shen R, Asa S, Ezzat S, Gonzalez-Cadavid NF. J Endocrinol. 2000 Dec; 167(3):417-28. https://www.ncbi.nlm.nih.gov/pubmed/11115768/
- Functional improvement of dystrophic muscle by myostatin blockade. Bogdanovich S, Krag TO, Barton ER, Morris LD, Whittemore LA, Ahima RS, Khurana TS. Nature. 2002 Nov 28; 420(6914):418-21. https://www.ncbi.nlm.nih.gov/pubmed/12459784/
- Ectopic expression of myostatin induces atrophy of adult skeletal muscle by decreasing muscle gene expression. Durieux AC, Amirouche A, Banzet S, Koulmann N, Bonnefoy R, Pasdeloup M, Mouret C, Bigard X, Peinnequin A, Freyssenet D. Endocrinology. 2007 Jul; 148(7):3140-7. https://www.ncbi.nlm.nih.gov/pubmed/17395701/
- Myostatin gene deletion prevents glucocorticoid-induced muscle atrophy. Gilson H, Schakman O, Combaret L, Lause P, Grobet L, Attaix D, Ketelslegers JM, Thissen JP. Endocrinology. 2007 Jan; 148(1):452-60. https://www.ncbi.nlm.nih.gov/pubmed/17038559/
- Liver-derived insulin-like growth factor I (IGF-I) is the principal source of IGF-I in blood but is not required for postnatal body growth in mice. Sjögren K, Liu JL, Blad K, Skrtic S, Vidal O, Wallenius V, LeRoith D, Törnell J, Isaksson OG, Jansson JO, Ohlsson C. Proc Natl Acad Sci U S A. 1999 Jun 8; 96(12):7088-92. https://www.ncbi.nlm.nih.gov/pubmed/10359843/
- Short-term growth hormone treatment does not increase muscle protein synthesis in experienced weight lifters. Yarasheski KE, Zachweija JJ, Angelopoulos TJ, Bier DM. J Appl Physiol (1985). 1993 Jun; 74(6):3073-6. https://www.ncbi.nlm.nih.gov/pubmed/8366011/
- PGC-1alpha regulation by exercise training and its influences on muscle function and insulin sensitivity. Lira VA, Benton CR, Yan Z, Bonen A. Am J Physiol Endocrinol Metab. 2010 Aug; 299(2):E145-61. https://www.ncbi.nlm.nih.gov/pubmed/20371735/
- Progressive effect of endurance training on metabolic adaptations in working skeletal muscle. Phillips SM, Green HJ, Tarnopolsky MA, Heigenhauser GJ, Grant SM. Am J Physiol. 1996 Feb; 270(2 Pt 1):E265-72. https://www.ncbi.nlm.nih.gov/pubmed/8779948/
- Exercise metabolism and the molecular regulation of skeletal muscle adaptation. Egan B, Zierath JR. Cell Metab. 2013 Feb 5; 17(2):162-84. https://www.ncbi.nlm.nih.gov/pubmed/23395166/
- Intracellular signaling specificity in skeletal muscle in response to different modes of exercise. Nader GA, Esser KA. J Appl Physiol (1985). 2001 May; 90(5):1936-42. http://jap.physiology.org/content/90/5/1936.long
- Mikines KJ, Sonne B, Farrell PA, Tronier B, and Galbo H. Effect of physical exercise on sensitivity and responsiveness to insulin in humans. Am J Physiol 254: E248–E259, 1988. https://www.ncbi.nlm.nih.gov/pubmed/3126668
- Gundersen K. Excitation-transcription coupling in skeletal muscle: the molecular pathways of exercise. Biol Rev Camb Philos Soc 86: 564–600, 2011. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3170710/
- Borst SE. Interventions for sarcopenia and muscle weakness in older people. Age Ageing 33: 548–555, 2004. https://www.ncbi.nlm.nih.gov/pubmed/15385272
- Fry AC. Sports Med. 2004;34(10):663-79. The role of resistance exercise intensity on muscle fibre adaptations. https://www.ncbi.nlm.nih.gov/pubmed/15335243
- PI3 kinase regulation of skeletal muscle hypertrophy and atrophy. Glass DJ. Curr Top Microbiol Immunol. 2010; 346():267-78. https://www.ncbi.nlm.nih.gov/pubmed/20593312/
- Autophagy: renovation of cells and tissues. Mizushima N, Komatsu M. Cell. 2011 Nov 11; 147(4):728-41. https://www.ncbi.nlm.nih.gov/pubmed/22078875/
- Physiological role of autophagy as an intracellular recycling system: with an emphasis on nutrient metabolism. Kuma A, Mizushima N. Semin Cell Dev Biol. 2010 Sep; 21(7):683-90. https://www.ncbi.nlm.nih.gov/pubmed/20223289/
- Protein breakdown in muscle wasting: role of autophagy-lysosome and ubiquitin-proteasome. Sandri M. Int J Biochem Cell Biol. 2013 Oct; 45(10):2121-9. https://www.ncbi.nlm.nih.gov/pubmed/23665154/
- FoxO3 controls autophagy in skeletal muscle in vivo. Mammucari C, Milan G, Romanello V, Masiero E, Rudolf R, Del Piccolo P, Burden SJ, Di Lisi R, Sandri C, Zhao J, Goldberg AL, Schiaffino S, Sandri M. Cell Metab. 2007 Dec; 6(6):458-71. https://www.ncbi.nlm.nih.gov/pubmed/18054315/
- Cellular and molecular mechanisms of muscle atrophy. Bonaldo P, Sandri M. Dis Model Mech. 2013 Jan; 6(1):25-39. https://www.ncbi.nlm.nih.gov/pubmed/23268536/
- Grumati P, Coletto L, Schiavinato A, Castagnaro S, Bertaggia E, Sandri M, and Bonaldo P. Physical exercise stimulates autophagy in normal skeletal muscles but is detrimental for collagen VI-deficient muscles. Autophagy 7: 1415–1423, 2011. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3288016/
- Ferraro E, Giammarioli AM, Chiandotto S, Spoletini I, Rosano G. Exercise-Induced Skeletal Muscle Remodeling and Metabolic Adaptation: Redox Signaling and Role of Autophagy. Antioxidants & Redox Signaling. 2014;21(1):154-176. doi:10.1089/ars.2013.5773. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4048572
- Abernethy PJ, Jürimäe J, Logan PA, Taylor AW, Thayer RE. Sports Med. 1994 Jan;17(1):22-38. Acute and chronic response of skeletal muscle to resistance exercise. https://www.ncbi.nlm.nih.gov/pubmed/8153497?dopt=Abstract
- Tesch PA. Med Sci Sports Exerc. 1988 Oct;20(5 Suppl):S132-4. Skeletal muscle adaptations consequent to long-term heavy resistance exercise. https://www.ncbi.nlm.nih.gov/pubmed/3057312
- Campos GE, Luecke TJ, Wendeln HK, Toma K, Hagerman FC, Murray TF, Ragg KE, Ratamess NA, Kraemer WJ, Staron RS. Eur J Appl Physiol. 2002 Nov;88(1-2):50-60. Epub 2002 Aug 15. Muscular adaptations in response to three different resistance-training regimens: specificity of repetition maximum training zones. https://www.ncbi.nlm.nih.gov/pubmed/12436270
- Functional properties of human muscle fibers after short-term resistance exercise training. Jeffrey J. Widrick, Julian E. Stelzer, Todd C. Shoepe, Dena P. Garner. American Journal of Physiology – Regulatory, Integrative and Comparative Physiology Aug 2002, 283 (2) R408-R416; DOI: 10.1152/ajpregu.00120.2002. http://ajpregu.physiology.org/content/283/2/R408.long
- Hather BM, Tesch PA, Buchanan P, Dudley GA. (1991) Influence of eccentric actions on skeletal muscle adaptations to resistance training. Acta Physiol Scand 143:177–185. https://www.ncbi.nlm.nih.gov/pubmed/1835816
- Journal of Applied Physiology Nov 1996, 81 (5) 2004-2012; Muscle fiber hypertrophy, hyperplasia, and capillary density in college men after resistance training. http://jap.physiology.org/content/81/5/2004
- Aragon AA, Schoenfeld BJ, Wildman R, et al. International society of sports nutrition position stand: diets and body composition. Journal of the International Society of Sports Nutrition. 2017;14:16. doi:10.1186/s12970-017-0174-y. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5470183/
- Hall KD. What is the required energy deficit per unit weight loss? Int J Obes. 2007;32:573–576. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2376744/
- Garthe I, Raastad T, Refsnes PE, Koivisto A, Sundgot-Borgen J. Effect of two different weight-loss rates on body composition and strength and power-related performance in elite athletes. Int J Sport Nutr Exerc Metab. 2011;21:97–104. https://www.ncbi.nlm.nih.gov/pubmed/21558571
- Hall KD. Body fat and fat-free mass inter-relationships: Forbes’s theory revisited. Br J Nutr. 2007;97:1059–1063. doi: 10.1017/S0007114507691946. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2376748/
- Mero AA, Huovinen H, Matintupa O, Hulmi JJ, Puurtinen R, Hohtari H, Karila T. Moderate energy restriction with high protein diet results in healthier outcome in women. J Int Soc Sports Nutr. 2010;7:4. doi: 10.1186/1550-2783-7-4. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2822830/
- Withers RT, Noell CJ, Whittingham NO, Chatterton BE, Schultz CG, Keeves JP. Body composition changes in elite male bodybuilders during preparation for competition. Aust J Sci Med Sport. 1997;29:11–16. https://www.ncbi.nlm.nih.gov/pubmed/9127683
- van der Ploeg GE, Brooks AG, Withers RT, Dollman J, Leaney F, Chatterton BE. Body composition changes in female bodybuilders during preparation for competition. Eur J Clin Nutr. 2001;55:268–277. doi: 10.1038/sj.ejcn.1601154. https://www.ncbi.nlm.nih.gov/pubmed/11360131
- Newton LE, Hunter GR, Bammon M, Roney RK. Changes in psychological state and self-reported diet during various phases of training in competitive bodybuilders. J Strength Cond Res. 1993;7:153–158.
- Forbes GB. Body fat content influences the body composition response to nutrition and exercise. Ann N Y Acad Sci. 2000;904:359–365. https://www.ncbi.nlm.nih.gov/pubmed/10865771
- Harvie MN, Pegington M, Mattson MP, Frystyk J, Dillon B, Evans G, Cuzick J, Jebb SA, Martin B, Cutler RG, Son TG, Maudsley S, Carlson OD, Egan JM, Flyvbjerg A, Howell A. The effects of intermittent or continuous energy restriction on weight loss and metabolic disease risk markers: a randomized trial in young overweight women. Int J Obes. 2011;35:714–727. doi: 10.1038/ijo.2010.171. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3017674/
- Soeters MR, Lammers NM, Dubbelhuis PF, Ackermans M, Jonkers-Schuitema CF, Fliers E, Sauerwein HP, Aerts JM, Serlie MJ. Intermittent fasting does not affect whole-body glucose, lipid, or protein metabolism. Am J Clin Nutr. 2009;90:1244–1251. doi: 10.3945/ajcn.2008.27327. https://www.ncbi.nlm.nih.gov/pubmed/19776143
- La Bounty PM, Campbell BI, Wilson J, Galvan E, Berardi J, Kleiner SM, Kreider RB, Stout JR, Ziegenfuss T, Spano M, Smith A, Antonio J. International Society of Sports Nutrition position stand: meal frequency. J Int Soc Sports Nutr. 2011;8:4. doi: 10.1186/1550-2783-8-4. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3070624/
- Compared with nibbling, neither gorging nor a morning fast affect short-term energy balance in obese patients in a chamber calorimeter. Taylor MA, Garrow JS. Int J Obes Relat Metab Disord. 2001 Apr; 25(4):519-28. https://www.ncbi.nlm.nih.gov/pubmed/11319656/
- Influence of the feeding frequency on nutrient utilization in man: consequences for energy metabolism. Verboeket-van de Venne WP, Westerterp KR. Eur J Clin Nutr. 1991 Mar; 45(3):161-9. https://www.ncbi.nlm.nih.gov/pubmed/1905998/
- Helms ER, Aragon AA, Fitschen PJ. Evidence-based recommendations for natural bodybuilding contest preparation: nutrition and supplementation. Journal of the International Society of Sports Nutrition. 2014;11:20. doi:10.1186/1550-2783-11-20. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4033492/
- Campbell B, Kreider R, Ziegenfuss T, La Bounty P, Roberts M, Burke D, et al. International Society of Sports Nutrition position stand: protein and exercise. J Int Soc Sports Nutr. 2007;4:8. doi: 10.1186/1550-2783-4-8. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2117006/
- Helms E, Zinn C, Rowlands D, Brown S. A systematic review of dietary protein during caloric restriction in resistance trained lean athletes: a case for higher intakes. Int J Sport Nutr Exerc Metab. 2014;24(2):127–38. doi: 10.1123/ijsnem.2013-0054. https://www.ncbi.nlm.nih.gov/pubmed/24092765
- Helms E, Aragon A, Fitschen P. Evidence-based recommendations for natural bodybuilding contest preparation: nutrition and supplementation. J Int Soc Sports Nutr. 2014;11:20. doi: 10.1186/1550-2783-11-20. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4033492/
- Bandegan A, Courtney-Martin G, Rafii M, Pencharz P, Lemon P. Indicator amino acid–derived estimate of dietary protein requirement for male bodybuilders on a non training day is several-fold greater than the current recommended dietary allowance. J Nutr. 2017;147(5):850-7. https://www.ncbi.nlm.nih.gov/pubmed/28179492
- Cermak NR, de PT, Groot LC S, WH van Loon LJ. Protein supplementation augments the adaptive response of skeletal muscle to resistance-type exercise training: a meta-analysis. Am J Clin Nutr. 2012;96(6):1454–64. doi: 10.3945/ajcn.112.037556. http://ajcn.nutrition.org/content/96/6/1454.long
- Phillips S, Van Loon L. Dietary protein for athletes: from requirements to optimum adaptation. J Sports Sci. 2011;29(Suppl 1):S29–38. doi: 10.1080/02640414.2011.619204. https://www.ncbi.nlm.nih.gov/pubmed/22150425
- Churchward-Venne T, Murphy C, Longland T, Phillips S. Role of protein and amino acids in promoting lean mass accretion with resistance exercise and attenuating lean mass loss during energy deficit in humans. Amino Acids. 2013;45(2):231–40. doi: 10.1007/s00726-013-1506-0. https://www.ncbi.nlm.nih.gov/pubmed/23645387
- Attarzadeh Hosseini S, Sardar M, Hejazi K, Farahati S. The effect of ramadan fasting and physical activity on body composition, serum osmolarity levels and some parameters of electrolytes in females. Int J Endocrinol Metab. 2013;11(2):88–94. doi: 10.5812/ijem.9602. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3693661/
- Norouzy A, Salehi M, Philippou E, Arabi H, Shiva F, Mehrnoosh S, Mohajeri SMR, Reza Mohajeri SA, Motaghedi Larijani A, Nematy M. Effect of fasting in Ramadan on body composition and nutritional intake: a prospective study. J Hum Nutr Diet. 2013;26(Suppl. 1):97–104. https://www.ncbi.nlm.nih.gov/pubmed/23679071
- Time-restricted feeding in young men performing resistance training: A randomized controlled trial. Tinsley GM, Forsse JS, Butler NK, Paoli A, Bane AA, La Bounty PM, Morgan GB, Grandjean PW. Eur J Sport Sci. 2017 Mar; 17(2):200-207. https://www.ncbi.nlm.nih.gov/pubmed/27550719/
- Effects of eight weeks of time-restricted feeding (16/8) on basal metabolism, maximal strength, body composition, inflammation, and cardiovascular risk factors in resistance-trained males. Moro T, Tinsley G, Bianco A, Marcolin G, Pacelli QF, Battaglia G, Palma A, Gentil P, Neri M, Paoli A. J Transl Med. 2016 Oct 13; 14(1):290. https://www.ncbi.nlm.nih.gov/pubmed/27737674/
- Do intermittent diets provide physiological benefits over continuous diets for weight loss? A systematic review of clinical trials. Seimon RV, Roekenes JA, Zibellini J, Zhu B, Gibson AA, Hills AP, Wood RE, King NA, Byrne NM, Sainsbury A. Mol Cell Endocrinol. 2015 Dec 15; 418 Pt 2():153-72. https://www.ncbi.nlm.nih.gov/pubmed/26384657/
- Kleiner SM, Bazzarre TL, Litchford MD. Metabolic profiles, diet, and health practices of championship male and female bodybuilders. J Am Diet Assoc. 1990;90:962–967. https://www.ncbi.nlm.nih.gov/pubmed/2365938
- Hickson JF Jr, Johnson TE, Lee W, Sidor RJ. Nutrition and the precontest preparations of a male bodybuilder. J Am Diet Assoc. 1990;90:264–267. https://www.ncbi.nlm.nih.gov/pubmed/2303663
- Andersen RE, Barlett SJ, Morgan GD, Brownell KD. Weight loss, psychological, and nutritional patterns in competitive male body builders. Int J Eat Disord. 1995 Jul;18(1):49-57. https://www.ncbi.nlm.nih.gov/pubmed/7670443
- Balon TW, Horowitz JF, Fitzsimmons KM. Effects of carbohydrate loading and weight-lifting on muscle girth. Int J Sport Nutr. 1992;2:328–334. https://www.ncbi.nlm.nih.gov/pubmed/1299502
- Costill DL, Cote R, Fink W. Muscle water and electrolytes following varied levels of dehydration in man. J Appl Physiol. 1976;40:6–11. https://www.ncbi.nlm.nih.gov/pubmed/1248983
- Maestu J, Eliakim A, Jurimae J, Valter I, Jurimae T. Anabolic and catabolic hormones and energy balance of the male bodybuilders during the preparation for the competition. J Strength Cond Res. 2010;24:1074–1081. doi: 10.1519/JSC.0b013e3181cb6fd3. https://www.ncbi.nlm.nih.gov/pubmed/20300017
- Bamman MM, Hunter GR, Newton LE, Roney RK, Khaled MA. Changes in body composition, diet, and strength of bodybuilders during the 12 weeks prior to competition. J Sports Med Phys Fitness. 1993;33:383–391. https://www.ncbi.nlm.nih.gov/pubmed/8035587
- Walberg-Rankin J, Edmonds CE, Gwazdauskas FC. Diet and weight changes of female bodybuilders before and after competition. Int J Sport Nutr. 1993;3:87–102. https://www.ncbi.nlm.nih.gov/pubmed/8499941
- Shephard RJ. Electrolyte manipulation in female body-builders. Br J Sports Med. 1994;28:60–61. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1332163/
- Aragon AA, Schoenfeld BJ, Wildman R, et al. International society of sports nutrition position stand: diets and body composition. Journal of the International Society of Sports Nutrition. 2017;14:16. doi:10.1186/s12970-017-0174-y. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5470183
- Joosen A, Westerterp K. Energy expenditure during overfeeding. Nutr Metab (Lond) 2006;3:25. doi: 10.1186/1743-7075-3-25. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1543621/
- Changes in energy expenditure resulting from altered body weight. Leibel RL, Rosenbaum M, Hirsch J. N Engl J Med. 1995 Mar 9; 332(10):621-8. https://www.ncbi.nlm.nih.gov/pubmed/7632212/
- Determinants of 24-hour energy expenditure in man. Methods and results using a respiratory chamber. Ravussin E, Lillioja S, Anderson TE, Christin L, Bogardus C. J Clin Invest. 1986 Dec; 78(6):1568-78. https://www.ncbi.nlm.nih.gov/pubmed/3782471/
- Joosen AM, Westerterp KR. Energy expenditure during overfeeding. Nutrition & Metabolism. 2006;3:25. doi:10.1186/1743-7075-3-25. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1543621/
- Amino Acids. 2013 Aug;45(2):231-40. doi: 10.1007/s00726-013-1506-0. Epub 2013 May 5. Role of protein and amino acids in promoting lean mass accretion with resistance exercise and attenuating lean mass loss during energy deficit in humans. https://www.ncbi.nlm.nih.gov/pubmed/23645387
- Dietary protein for athletes: from requirements to optimum adaptation. Phillips SM, Van Loon LJ. J Sports Sci. 2011; 29 Suppl 1():S29-38. https://www.ncbi.nlm.nih.gov/pubmed/22150425/
- Contemporary issues in protein requirements and consumption for resistance trained athletes. Wilson J, Wilson GJ. J Int Soc Sports Nutr. 2006 Jun 5; 3():7-27. https://www.ncbi.nlm.nih.gov/pubmed/18500966/
- Comparison of body composition, exercise and nutritional profiles of female and male body builders at competition. Sandoval WM, Heyward VH, Lyons TM. J Sports Med Phys Fitness. 1989 Mar; 29(1):63-70. https://www.ncbi.nlm.nih.gov/pubmed/2770270/
- A systematic review of dietary protein during caloric restriction in resistance trained lean athletes: a case for higher intakes. Helms ER, Zinn C, Rowlands DS, Brown SR. Int J Sport Nutr Exerc Metab. 2014 Apr; 24(2):127-38. https://www.ncbi.nlm.nih.gov/pubmed/24092765/
- Muscle substrate utilization and lactate production. MacDougall JD, Ray S, Sale DG, McCartney N, Lee P, Garner S. Can J Appl Physiol. 1999 Jun; 24(3):209-15. https://www.ncbi.nlm.nih.gov/pubmed/10364416/
- Nutrition guidelines for strength sports: sprinting, weightlifting, throwing events, and bodybuilding. Slater G, Phillips SM. J Sports Sci. 2011; 29 Suppl 1():S67-77. https://www.ncbi.nlm.nih.gov/pubmed/21660839/
- A reduced ratio of dietary carbohydrate to protein improves body composition and blood lipid profiles during weight loss in adult women. Layman DK, Boileau RA, Erickson DJ, Painter JE, Shiue H, Sather C, Christou DD. J Nutr. 2003 Feb; 133(2):411-7. https://www.ncbi.nlm.nih.gov/pubmed/12566476/
- Energy expenditure, satiety, and plasma ghrelin, glucagon-like peptide 1, and peptide tyrosine-tyrosine concentrations following a single high-protein lunch. Smeets AJ, Soenen S, Luscombe-Marsh ND, Ueland Ø, Westerterp-Plantenga MS. J Nutr. 2008 Apr; 138(4):698-702. https://www.ncbi.nlm.nih.gov/pubmed/18356323/
- Low-carbohydrate diets and performance. Cook CM, Haub MD. Curr Sports Med Rep. 2007 Jul; 6(4):225-9. https://www.ncbi.nlm.nih.gov/pubmed/17617997/
- Anabolic and catabolic hormones and energy balance of the male bodybuilders during the preparation for the competition. Mäestu J, Eliakim A, Jürimäe J, Valter I, Jürimäe T. J Strength Cond Res. 2010 Apr; 24(4):1074-81. https://www.ncbi.nlm.nih.gov/pubmed/20300017/
- Pituitary–gonadal axis during prolonged total starvation in obese men. Suryanarayana BV, Kent JR, Meister L, Parlow AF. Am J Clin Nutr. 1969 Jun; 22(6):767-70. https://www.ncbi.nlm.nih.gov/pubmed/5789477/
- Moderate energy restriction with high protein diet results in healthier outcome in women. Mero AA, Huovinen H, Matintupa O, Hulmi JJ, Puurtinen R, Hohtari H, Karila TA. J Int Soc Sports Nutr. 2010 Jan 25; 7(1):4. https://www.ncbi.nlm.nih.gov/pubmed/20205751/
- Decrease of serum total and free testosterone during a low-fat high-fibre diet. Hämäläinen EK, Adlercreutz H, Puska P, Pietinen P. J Steroid Biochem. 1983 Mar; 18(3):369-70. https://www.ncbi.nlm.nih.gov/pubmed/6298507/
- Diet and serum sex hormones in healthy men. Hämäläinen E, Adlercreutz H, Puska P, Pietinen P. J Steroid Biochem. 1984 Jan; 20(1):459-64. https://www.ncbi.nlm.nih.gov/pubmed/6538617/
- Bird SP. Strength nutrition: maximizing your anabolic potential. Strength Cond J. 2010;32:80–86. doi: 10.1519/SSC.0b013e3181d5284e.
- Macronutrient content of a hypoenergy diet affects nitrogen retention and muscle function in weight lifters. Walberg JL, Leidy MK, Sturgill DJ, Hinkle DE, Ritchey SJ, Sebolt DR. Int J Sports Med. 1988 Aug; 9(4):261-6. https://www.ncbi.nlm.nih.gov/pubmed/3182156/
- Macronutrient considerations for the sport of bodybuilding. Lambert CP, Frank LL, Evans WJ. Sports Med. 2004; 34(5):317-27. https://www.ncbi.nlm.nih.gov/pubmed/15107010/
- A perspective on fat intake in athletes. Pendergast DR, Leddy JJ, Venkatraman JT. J Am Coll Nutr. 2000 Jun; 19(3):345-50. https://www.ncbi.nlm.nih.gov/pubmed/10872896/
- International Society of Sports Nutrition position stand: nutrient timing. Kerksick C, Harvey T, Stout J, Campbell B, Wilborn C, Kreider R, Kalman D, Ziegenfuss T, Lopez H, Landis J, Ivy JL, Antonio J. J Int Soc Sports Nutr. 2008 Oct 3; 5():17. https://www.ncbi.nlm.nih.gov/pubmed/18834505/
- Aragon AA, Schoenfeld BJ. Nutrient timing revisited: is there a post-exercise anabolic window? J Int Soc Sports Nutr. 2013;10:5. doi: 10.1186/1550-2783-10-5. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3577439/
- National Center for Complementary and Integrative Health. Bodybuilding. https://nccih.nih.gov/health/bodybuilding
- http://ocbonline.com/
- Nutrient intake, body fat, and lipid profiles of competitive male and female bodybuilders. Bazzarre TL, Kleiner SM, Litchford MD. J Am Coll Nutr. 1990 Apr; 9(2):136-42. https://www.ncbi.nlm.nih.gov/pubmed/2338462/
- Nutritional status of nationally ranked elite bodybuilders. Kleiner SM, Bazzarre TL, Ainsworth BE. Int J Sport Nutr. 1994 Mar; 4(1):54-69. https://www.ncbi.nlm.nih.gov/pubmed/8167655/
- Food selection patterns of bodybuilders. Sandoval WM, Heyward VH. Int J Sport Nutr. 1991 Mar; 1(1):61-8. https://www.ncbi.nlm.nih.gov/pubmed/1844403/
- Improved cycling time-trial performance after ingestion of a caffeine energy drink. Ivy JL, Kammer L, Ding Z, Wang B, Bernard JR, Liao YH, Hwang J. Int J Sport Nutr Exerc Metab. 2009 Feb; 19(1):61-78. https://www.ncbi.nlm.nih.gov/pubmed/19403954/
- The effects of caffeine ingestion on time trial cycling performance. McNaughton LR, Lovell RJ, Siegler J, Midgley AW, Moore L, Bentley DJ. Int J Sports Physiol Perform. 2008 Jun; 3(2):157-63. https://www.ncbi.nlm.nih.gov/pubmed/19208924/
- Carr A, Dawson B, Schneiker K, Goodman C, Lay B. Effect of caffeine supplementation on repeated sprint running performance. J Sports Med Phys Fitness. 2008;48:472–478. https://www.ncbi.nlm.nih.gov/pubmed/18997650
- Glaister M, Howatson G, Abraham CS, Lockey RA, Goodwin JE, Foley P, McInnes G. Caffeine supplementation and multiple sprint running performance. Med Sci Sports Exerc. 2008;40:1835–1840. doi: 10.1249/MSS.0b013e31817a8ad2. https://www.ncbi.nlm.nih.gov/pubmed/18799995
- Green JM, Wickwire PJ, McLester JR, Gendle S, Hudson G, Pritchett RC, Laurent CM. Effects of caffeine on repetitions to failure and ratings of perceived exertion during resistance training. Int J Sports Physiol Perform. 2007;2:250–259. https://www.ncbi.nlm.nih.gov/pubmed/19168925
- Woolf K, Bidwell WK, Carlson AG. The effect of caffeine as an ergogenic aid in anaerobic exercise. Int J Sport Nutr Exerc Metab. 2008;18:412–429. https://www.ncbi.nlm.nih.gov/pubmed/18708685
- Duncan MJ, Oxford SW. The effect of caffeine ingestion on mood state and bench press performance to failure. J Strength Cond Res. 2011;25:178–185. doi: 10.1519/JSC.0b013e318201bddb. https://www.ncbi.nlm.nih.gov/pubmed/21157384
- Williams AD, Cribb PJ, Cooke MB, Hayes A. The effect of ephedra and caffeine on maximal strength and power in resistance-trained athletes. J Strength Cond Res. 2008;22:464–470. doi: 10.1519/JSC.0b013e3181660320. https://www.ncbi.nlm.nih.gov/pubmed/18550961
- Hendrix CR, Housh TJ, Mielke M, Zuniga JM, Camic CL, Johnson GO, Schmidt RJ, Housh DJ. Acute effects of a caffeine-containing supplement on bench press and leg extension strength and time to exhaustion during cycle ergometry. J Strength Cond Res. 2010;24:859–865. doi: 10.1519/JSC.0b013e3181ae7976. https://www.ncbi.nlm.nih.gov/pubmed/19834348
- Nawrot P, Jordan S, Eastwood J, Rotstein J, Hugenholtz A, Feeley M. Effects of caffeine on human health. Food Addit Contam. 2003;20:1–30. https://www.ncbi.nlm.nih.gov/pubmed/12519715
- Tarnopolsky MA, Atkinson SA, MacDougall JD, Sale DG, Sutton JR. Physiological responses to caffeine during endurance running in habitual caffeine users. Med Sci Sports Exerc. 1989;21:418–424. https://www.ncbi.nlm.nih.gov/pubmed/2674593
- Artioli GG, Gualano B, Smith A, Stout J, Lancha AH Jr. Role of beta-alanine supplementation on muscle carnosine and exercise performance. Med Sci Sports Exerc. 2010;42:1162–1173. https://www.ncbi.nlm.nih.gov/pubmed/20479615
- Harris RC, Tallon MJ, Dunnett M, Boobis L, Coakley J, Kim HJ, Fallowfield JL, Hill CA, Sale C, Wise JA. The absorption of orally supplied beta-alanine and its effect on muscle carnosine synthesis in human vastus lateralis. Amino Acids. 2006;30:279–289. doi: 10.1007/s00726-006-0299-9. https://www.ncbi.nlm.nih.gov/pubmed/16554972
- Hobson RM, Saunders B, Ball G, Harris RC, Sale C. Effects of beta-alanine supplementation on exercise performance: a meta-analysis. . Amino Acids. 2012;43:25–37. doi: 10.1007/s00726-011-1200-z. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3374095/
- Lu P, Xu W, Sturman JA. Dietary beta-alanine results in taurine depletion and cerebellar damage in adult cats. J Neurosci Res. 1996;43:112–119. doi: 10.1002/jnr.490430115. https://www.ncbi.nlm.nih.gov/pubmed/8838582
- Smith HJ, Mukerji P, Tisdale MJ. Attenuation of proteasome-induced proteolysis in skeletal muscle by {beta}-hydroxy-{beta}-methylbutyrate in cancer-induced muscle loss. Cancer Res. 2005;65:277–283. https://www.ncbi.nlm.nih.gov/pubmed/15665304
- Eley HL, Russell ST, Baxter JH, Mukerji P, Tisdale MJ. Signaling pathways initiated by beta-hydroxy-beta-methylbutyrate to attenuate the depression of protein synthesis in skeletal muscle in response to cachectic stimuli. Am J Physiol Endocrinol Metab. 2007;293:E923–E931. doi: 10.1152/ajpendo.00314.2007. https://www.ncbi.nlm.nih.gov/pubmed/17609254
- Rathmacher JA, Nissen S, Panton L, Clark RH, Eubanks May P, Barber AE, D’Olimpio J, Abumrad NN. Supplementation with a combination of beta-hydroxy-beta-methylbutyrate (HMB), arginine, and glutamine is safe and could improve hematological parameters. JPEN J Parenter Enteral Nutr. 2004;28:65–75. doi: 10.1177/014860710402800265. https://www.ncbi.nlm.nih.gov/pubmed/15080599
- Gallagher PM, Carrithers JA, Godard MP, Schulze KE, Trappe SW. Beta-hydroxy-beta-methylbutyrate ingestion, part II: effects on hematology, hepatic and renal function. Med Sci Sports Exerc. 2000;32:2116–2119. doi: 10.1097/00005768-200012000-00023. https://www.ncbi.nlm.nih.gov/pubmed/11128860
- Fitschen PJ, Wilson GJ, Wilson JM, Wilund KR. Efficacy of beta-hydroxy-beta-methylbutyrate supplementation in elderly and clinical populations. Nutrition. 2013;29:29–36. doi: 10.1016/j.nut.2012.05.005. https://www.ncbi.nlm.nih.gov/pubmed/23085015
- Wilson GJ, Wilson JM, Manninen AH. Effects of beta-hydroxy-beta-methylbutyrate (HMB) on exercise performance and body composition across varying levels of age, sex, and training experience: a review. Nutr Metab (Lond) 2008;5:1. doi: 10.1186/1743-7075-5-1. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2245953/
- Wilson J, Fitschen P, Campbell B, Wilson G, Zanchi N, Taylor L, Wilborn C, Kalman D, Stout J, Hoffman J, Ziegenfuss T, Lopez H, Kreider R, Smith-Ryan A, Antonio J. International Society of Sports Nutrition Position Stand: beta-hydroxy-beta-methylbutyrate (HMB) J Int Soc Sports Nutr. 2013;10:6. doi: 10.1186/1550-2783-10-6. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3568064/
- University of Maryland Medical Center. Creatine. http://www.umm.edu/health/medical/altmed/supplement/creatine
- Cooper R, Naclerio F, Allgrove J, Jimenez A. Creatine supplementation with specific view to exercise/sports performance: an update. Journal of the International Society of Sports Nutrition. 2012;9:33. doi:10.1186/1550-2783-9-33. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3407788/
- Clinical pharmacology of the dietary supplement creatine monohydrate. Persky AM, Brazeau GA. Pharmacol Rev. 2001 Jun; 53(2):161-76. https://www.ncbi.nlm.nih.gov/pubmed/11356982/
- Brunzel NA. Renal function: Nonprotein nitrogen compounds, function tests, and renal disease. In: Scardiglia J, Brown M, McCullough K, Davis K, editor. Clinical Chemistry. McGraw-Hill: New York, NY; 2003. pp. 373–399.
- Potential ergogenic effects of arginine and creatine supplementation. Paddon-Jones D, Børsheim E, Wolfe RR. J Nutr. 2004 Oct; 134(10 Suppl):2888S-2894S; discussion 2895S. http://jn.nutrition.org/content/134/10/2888S.long
- Creatine in humans with special reference to creatine supplementation. Balsom PD, Söderlund K, Ekblom B. Sports Med. 1994 Oct; 18(4):268-80. https://www.ncbi.nlm.nih.gov/pubmed/7817065/
- Muscle creatine loading in men. Hultman E, Söderlund K, Timmons JA, Cederblad G, Greenhaff PL. J Appl Physiol (1985). 1996 Jul; 81(1):232-7. https://www.ncbi.nlm.nih.gov/pubmed/8828669/
- Buford TW, Kreider RB, Stout JR, et al. International Society of Sports Nutrition position stand: creatine supplementation and exercise. Journal of the International Society of Sports Nutrition. 2007;4:6. doi:10.1186/1550-2783-4-6. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2048496/
- The metabolic burden of creatine synthesis. Brosnan JT, da Silva RP, Brosnan ME. Amino Acids. 2011 May; 40(5):1325-31. https://www.ncbi.nlm.nih.gov/pubmed/21387089/
- Hultman E, Bergstrom J, Spreit L, Soderlund K. Energy metabolism and fatigue. In: Taylor A, Gollnick PD, Green H, editor. Biochemistry of Exercise VII. Human Kinetics: Champaign, IL; 1990. pp. 73–92.
- Greenhaff P. The nutritional biochemistry of creatine. J Nutrit Biochem. 1997;11:610–618. doi: 10.1016/S0955-2863(97)00116-2.
- Creatine supplementation and exercise performance: an update. Williams MH, Branch JD. J Am Coll Nutr. 1998 Jun; 17(3):216-34. https://www.ncbi.nlm.nih.gov/pubmed/9627907/
- Williams MH, Kreider R, Branch JD. Creatine: The power supplement. Champaign, IL: Human Kinetics Publishers; 1999. p. 252.
- Effect of creatine supplementation and resistance-exercise training on muscle insulin-like growth factor in young adults. Burke DG, Candow DG, Chilibeck PD, MacNeil LG, Roy BD, Tarnopolsky MA, Ziegenfuss T. Int J Sport Nutr Exerc Metab. 2008 Aug; 18(4):389-98. https://www.ncbi.nlm.nih.gov/pubmed/18708688/
- Chanutin A. The fate of creatine when administered to man. J Biol Chem. 1926;67:29–34.
- Buford T, Kreider R, Stout J, Greenwood M, Campbell B, Spano M, Ziegenfuss T, Lopez H, Landis J, Antonio J. International Society of Sports Nutrition position stand: creatine supplementation and exercise. J Int Soc Sports Nutr. 2007;4:6. doi: 10.1186/1550-2783-4-6. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2048496/
- American College of Sport Medicine. Round Table, the physiological and health effects of oral creatine supplementation. Med Sci Sports Exc. 2000;32:706–717. https://www.ncbi.nlm.nih.gov/pubmed/10731017
- Branch JD. Effects of creatine supplementation on body composition and performace: a meta análisis. Int J Sports Nutr Exerc Metabol. 2003;13:I198-122. https://www.ncbi.nlm.nih.gov/pubmed/12945830
- Rawson ES, Volek JS. Effects of creatine supplementation and resistance training on muscle strength and weightlifting performance. J Strength Cond Res. 2003;17:822–831. https://www.ncbi.nlm.nih.gov/pubmed/14636102
- Volek JS, Kraemer WJ. Creatine suplemetation: its effects on human muscular performance and body composition. J Strength Cond Res. 1996;10:200–210.
- Syrotuik DG, Bell GJ. Acute creatine monohydrate supplementation: a descriptive physiological profile of responders vs. nonresponders. J Strength Cond Res. 2004;18:610–617. https://www.ncbi.nlm.nih.gov/pubmed/15320650
- Greenhaff PL, Bodin K, Soderlund K, Hultman E. Effect of oral creatine supplementation on skeletal muscle phosphocreatine resynthesis. Am J Physiol. 1994;266:E725–E730. https://www.ncbi.nlm.nih.gov/pubmed/8203511
- Casey A, Greenhaff P. Does dietary creatine supplementation play a role in skeletal muscle metabolism and performance? Am J Clin Nutr. 2000;72:607S–617S. https://www.ncbi.nlm.nih.gov/pubmed/10919967
- Cribb PJ, Williams AD, Hayes A. A creatine-protein-carbohydrate supplement enhances responses to resistance training. Med Sci Sports Exerc. 2007;39:1960–1968. doi: 10.1249/mss.0b013e31814fb52a. https://www.ncbi.nlm.nih.gov/pubmed/17986903
- Parise G, Mihic S, MacLennan D, Yarasheski KE, Tarnopolsky MA. Effects of acute creatine monohydrate supplementation on leucine kinetics and mixed-muscle protein synthesis. J Appl Physiol. 2001;91:1041–1047. https://www.ncbi.nlm.nih.gov/pubmed/11509496
- Louis M, Poortmans JR, Francaux M, Hultman E, Berre J, Boisseau N, Young VR, Smith K, Meier-Augenstein W, Babraj JA. et al. Creatine supplementation has no effect on human muscle protein turnover at rest in the postabsorptive or fed states. Am J Physiol Endocrinol Metab. 2003;284:E764–E770. https://www.ncbi.nlm.nih.gov/pubmed/12475751
- Deldicque L, Atherton P, Patel R, Theisen D, Nielens H, Rennie M, Francaux M. Effects of resistance exercise with and without creatine supplementation on gene expression and cell signaling in human skeletal muscle. J Appl Physiol. 2008;104:371–378. https://www.ncbi.nlm.nih.gov/pubmed/18048590
- Burke DG, Candow DG, Chilibeck PD, MacNeil LG, Roy BD, Tarnopolsky MA, Ziegenfuss T. Effect of creatine supplementation and resistance-exercise training on muscle insulin-like growth factor in young adults. Int J Sport Nutr Exerc Metab. 2008;18:389–398. https://www.ncbi.nlm.nih.gov/pubmed/18708688
- Harp JB, Goldstein S, Phillips LS. Nutrition and somatomedin. XXIII. Molecular regulation of IGF-I by amino acid availability in cultured hepatocytes. Diabetes. 1991;40:95–101. doi: 10.2337/diabetes.40.1.95. https://www.ncbi.nlm.nih.gov/pubmed/1901809
- Op ‘t Eijnde B, Urso B, Richter EA, Greenhaff PL, Hespel P. Effect of oral creatine supplementation on human muscle GLUT4 protein content after immobilization. Diabetes. 2001;50:18–23. doi: 10.2337/diabetes.50.1.18. https://www.ncbi.nlm.nih.gov/pubmed/11147785
- Cooke MB, Rybalka E, Williams AD, Cribb PJ, Hayes A. Creatine supplementation enhances muscle force recovery after eccentrically-induced muscle damage in healthy individuals. J Int Soc Sports Nutr. 2009;6:13. doi: 10.1186/1550-2783-6-13. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2697134/
- Lawler JM, Barnes WS, Wu G, Song W, Demaree S. Direct antioxidant properties of creatine. Biochem Biophys Res Commun. 2002;290:47–52. doi: 10.1006/bbrc.2001.6164. https://www.ncbi.nlm.nih.gov/pubmed/11779131
- Sestili P, Martinelli C, Bravi G, Piccoli G, Curci R, Battistelli M, Falcieri E, Agostini D, Gioacchini AM, Stocchi V. Creatine supplementation affords cytoprotection in oxidatively injured cultured mammalian cells via direct antioxidant activity. Free Radic Biol Med. 2006;40:837–849. doi: 10.1016/j.freeradbiomed.2005.10.035. https://www.ncbi.nlm.nih.gov/pubmed/16520236
- Rahimi R. Creatine supplementation decreases oxidative DNA damage and lipid peroxidation induced by a single bout of resistance exercise. J Strength Cond Res. 2011;25:3448–3455. doi: 10.1519/JSC.0b013e3182162f2b. https://www.ncbi.nlm.nih.gov/pubmed/22080314
- Hespel P, Derave W. Ergogenic effects of creatine in sports and rehabilitation. Subcell Biochem. 2007;46:245–259. doi: 10.1021/bi061646s. https://www.ncbi.nlm.nih.gov/pubmed/18652080
- Nelson A, Arnall D, Kokkonen J, Day R, Evans J. Muscle glycogen supercompensation is enhanced by prior creatine supplementation. Med Sci Sports Exerc. 2001;33:1096–1100. https://www.ncbi.nlm.nih.gov/pubmed/11445755
- Sewell D, Robinson T, Greenhaff P. Creatine supplementation does not affect human skeletal muscle glycogen content in the absence of prior exercise. J Appl Physiol. 2008;104:508–512. https://www.ncbi.nlm.nih.gov/pubmed/18032580
- Hickner R, Dyck D, Sklar J, Hatley H, Byrd P. Effect of 28 days of creatine ingestion on muscle metabolism and performance of a simulated cycling road race. J Int Soc Sports Nutr. 2010;7:26. doi: 10.1186/1550-2783-7-26. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2909923/
- Branch J. Effect of creatine supplementation on body composition and performance: a meta-analysis. Int J Sport Nutr Exerc Metab. 2003;13:198–226. https://www.ncbi.nlm.nih.gov/pubmed/12945830
- Chwalbiñska-Moneta J. Effect of creatine supplementation on aerobic performance and anaerobic capacity in elite rowers in the course of endurance training. Int J Sport Nutr Exerc Metab. 2003;13:173–183. https://www.ncbi.nlm.nih.gov/pubmed/12945828
- Graef J, Smith A, Kendall K, Fukuda D, Moon J, Beck T, Cramer J, Stout J. The effects of four weeks of creatine supplementation and high-intensity interval training on cardiorespiratory fitness: a randomized controlled trial. J Int Soc Sports Nutr. 2009;6:18. doi: 10.1186/1550-2783-6-18. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2780977/
- Thompson C, Kemp G, Sanderson A, Dixon R, Styles P, Taylor D, Radda G. Effect of creatine on aerobic and anaerobic metabolism in skeletal muscle in swimmers. Br J Sports Med. 1996;30:222–225. doi: 10.1136/bjsm.30.3.222. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1332335/
- Volek J, Duncan N, Mazzetti S, Staron R, Putukian M, Gómez A, Pearson D, Fink W, Kraemer W. Performance and muscle fiber adaptations to creatine supplementation and heavy resistance training. Med Sci Sports Exerc. 1999;31:1147–1156. doi: 10.1097/00005768-199908000-00011. https://www.ncbi.nlm.nih.gov/pubmed/10449017
- Volek J, Rawson E. Scientific basis and practical aspects of creatine supplementation for athletes. Nutrition. 2004;20:609–614. doi: 10.1016/j.nut.2004.04.014. https://www.ncbi.nlm.nih.gov/pubmed/15212742
- van Loon L, Oosterlaar A, Hartgens F, Hesselink M, Snow R, Wagenmakers A. Effects of creatine loading and prolonged creatine supplementation on body composition, fuel selection, sprint and endurance performance in humans. Clin Sci (Lond) 2003;104:153–162. doi: 10.1042/CS20020159. https://www.ncbi.nlm.nih.gov/pubmed/12546637
- Kreider RB. Effects of creatine supplementation on performance and training adaptations. Mol Cell Biochem. 2003;244:89–94. doi: 10.1023/A:1022465203458. https://www.ncbi.nlm.nih.gov/pubmed/12701815
- Jakobi J, Rice C, Curtin S, Marsh G. Contractile properties, fatigue and recovery are not influenced by short-term creatine supplementation in human muscle. Exp Physiol. 2000;85:451–460. doi: 10.1017/S0958067000020212. https://www.ncbi.nlm.nih.gov/pubmed/10918084
- Bemben MG, Witten MS, Carter JM, Eliot KA, Knehans AW, Bemben DA. The effects of supplementation with creatine and protein on muscle strength following a traditional resistance training program in middle-aged and older men. J Nutr Health Aging. 2010;14:155–159. doi: 10.1007/s12603-009-0124-8. https://www.ncbi.nlm.nih.gov/pubmed/20126965
- Safdar A, Yardley N, Snow R, Melov S, Tarnopolsky M. Global and targeted gene expression and protein content in skeletal muscle of young men following short-term creatine monohydrate supplementation. Physiol Genomics. 2008;32:219–228. https://www.ncbi.nlm.nih.gov/pubmed/17957000
- Saremi A, Gharakhanloo R, Sharghi S, Gharaati M, Larijani B, Omidfar K. Effects of oral creatine and resistance training on serum myostatin and GASP-1. Mol Cell Endocrinol. 2010;317:25–30. doi: 10.1016/j.mce.2009.12.019. https://www.ncbi.nlm.nih.gov/pubmed/20026378
- Bazzucchi I, Felici F, Sacchetti M. Effect of short-term creatine supplementation on neuromuscular function. Med Sci Sports Exerc. 2009;41:1934–41. doi: 10.1249/MSS.0b013e3181a2c05c. https://www.ncbi.nlm.nih.gov/pubmed/19727018
- Sculthorpe N, Grace F, Jones P, Fletcher I. The effect of short-term creatine loading on active range of movement. Appl Physiol Nutr Metab. 2010;35:507–511. doi: 10.1139/H10-036. https://www.ncbi.nlm.nih.gov/pubmed/20725117
- Hile A, Anderson J, Fiala K, Stevenson J, Casa D, Maresh C. Creatine supplementation and anterior compartment pressure during exercise in the heat in dehydrated men. J Athl Train. 2006;41:30–35. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1421498/
- Metzl JD, Small E, Levine SR, Gershel JC. Creatine use among young athletes. Pediatrics. 2001;108:421–425. doi: 10.1542/peds.108.2.421. https://www.ncbi.nlm.nih.gov/pubmed/11483809
- Evans MW, Ndetan H, Perko M, Williams R, Walker C. Dietary supplement use by children and adolescents in the United States to Enhance sport performance: results of the national health interview survey. J Prim Prev. 2012;33:3–12. doi: 10.1007/s10935-012-0261-4. https://www.ncbi.nlm.nih.gov/pubmed/22297456
- Unnithan VB, Veehof SH, Vella CA, Kern M. Is there a physiologic basis for creatine use in children and adolescents? J Strength Cond Res. 2001;15:524–528. https://www.ncbi.nlm.nih.gov/pubmed/11726268
- Yoshizumi W, Tsourounis C. Effects of creatine supplementation on renal function. J Herb Pharmacother. 2004;4:1–7. https://www.ncbi.nlm.nih.gov/pubmed/15273072
- Thorsteinsdottir B, Grande J, Garovic V. Acute renal failure in a young weight lifter taking multiple food supplements, including creatine monohydrate. J Ren Nutr. 2006;16:341–345. doi: 10.1053/j.jrn.2006.04.025. https://www.ncbi.nlm.nih.gov/pubmed/17046619
- Bizzarini E, De Angelis L. Is the use of oral creatine supplementation safe? J Sports Med Phys Fitness. 2004;44:411–416. https://www.ncbi.nlm.nih.gov/pubmed/15758854
- Poortmans J, Francaux M. Adverse effects of creatine supplementation: fact or fiction? Sports Med. 2000;30:155–170. doi: 10.2165/00007256-200030030-00002. https://www.ncbi.nlm.nih.gov/pubmed/10999421
- Pline K, Smith C. The effect of creatine intake on renal function. Ann Pharmacother. 2005;39:1093–1096. doi: 10.1345/aph.1E628. https://www.ncbi.nlm.nih.gov/pubmed/15886291
- Kim HJ, Kim CK, Carpentier A, Poortmans JR. Studies on the safety of creatine supplementation. Amino Acids. 2011;40:1409–1418. doi: 10.1007/s00726-011-0878-2. https://www.ncbi.nlm.nih.gov/pubmed/21399917
- Sale C, Harris RC, Florance J, Kumps A, Sanvura R, Poortmans JR. Urinary creatine and methylamine excretion following 4 x 5 g x day(-1) or 20 x 1 g x day(-1) of creatine monohydrate for 5 days. J Sports Sci. 2009;27:759–766. doi: 10.1080/02640410902838237. https://www.ncbi.nlm.nih.gov/pubmed/19437189
- Tarnopolsky M, Zimmer A, Paikin J, Safdar A, Aboud A, Pearce E, Roy B, Doherty T. Creatine monohydrate and conjugated linoleic acid improve strength and body composition following resistance exercise in older adults. PLoS One. 2007;2:e991. doi: 10.1371/journal.pone.0000991. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1994592/
- Cornelissen VA, Defoor JG, Stevens A, Schepers D, Hespel P, Decramer M, Mortelmans L, Dobbels F, Vanhaecke J, Fagard RH, Vanhees L. Effect of creatine supplementation as a potential adjuvant therapy to exercise training in cardiac patients: a randomized controlled trial. Clin Rehabil. 2010;24:988–999. doi: 10.1177/0269215510367995. https://www.ncbi.nlm.nih.gov/pubmed/20576665
- Schilling B, Stone M, Utter A, Kearney J, Johnson M, Coglianese R, Smith L, O’Bryant H, Fry A, Starks M. et al. Creatine supplementation and health variables: a retrospective study. Med Sci Sports Exerc. 2001;33:183–188. https://www.ncbi.nlm.nih.gov/pubmed/11224803
- Dalbo V, Roberts M, Stout J, Kerksick C. Putting to rest the myth of creatine supplementation leading to muscle cramps and dehydration. Br J Sports Med. 2008;42:567–573. doi: 10.1136/bjsm.2007.042473. https://www.ncbi.nlm.nih.gov/pubmed/18184753
- Watson G, Casa D, Fiala K, Hile A, Roti M, Healey J, Armstrong L, Maresh C. Creatine use and exercise heat tolerance in dehydrated men. J Athl Train. 2006;41:18–29. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1421496/
- Lopez R, Casa D, McDermott B, Ganio M, Armstrong L, Maresh C. Does creatine supplementation hinder exercise heat tolerance or hydration status? A systematic review with meta-analyses. J Athl Train. 2009;44:215–223. doi: 10.4085/1062-6050-44.2.215. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2657025/
- Hadjicharalambous M, Kilduff L, Pitsiladis Y. Brain serotonin and dopamine modulators, perceptual responses and endurance performance during exercise in the heat following creatine supplementation. J Int Soc Sports Nutr. 2008;5:14. doi: 10.1186/1550-2783-5-14. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2570654/
- Persky A, Brazeau G. Clinical pharmacology of the dietary supplement creatine monohydrate. Pharmacol Rev. 2001;53:161–176. https://www.ncbi.nlm.nih.gov/pubmed/11356982
- Dempsey R, Mazzone M, Meurer L. Does oral creatine supplementation improve strength? A meta-analysis. J Fam Pract. 2002;51:945–951. https://www.ncbi.nlm.nih.gov/pubmed/12485548
- Ganguly S, Jayappa S, Dash AK. Evaluation of the stability of creatine in solution prepared from effervescent creatine formulations. AAPS PharmSciTech. 2003;4:E25. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2750587/
- Jäger R, Harris RC, Purpura M, Francaux M. Comparison of new forms of creatine in raising plasma creatine levels. J Int Soc Sports Nutr. 2007;4:17. doi: 10.1186/1550-2783-4-17. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2206055/
- Jäger R, Metzger J, Lautmann K, Shushakov V, Purpura M, Geiss K, Maassen N. The effects of creatine pyruvate and creatine citrate on performance during high intensity exercise. J Int Soc Sports Nutr. 2008;5:4. doi: 10.1186/1550-2783-5-4. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2276184/
- Jäger R, Purpura M, Shao A, Inoue T, Kreider RB. Analysis of the efficacy, safety, and regulatory status of novel forms of creatine. Amino Acids. 2011;40:1369–83. doi: 10.1007/s00726-011-0874-6. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3080578/
- Gufford BT, Sriraghavan K, Miller N, Miller D, Gu X, Vennerstrom J, Robinson D. Physicochemical characterization of creatine N-methylguanidinium salts. Journal of Dietary Supplements. 2010;7:240–252. doi: 10.3109/19390211.2010.491507. https://www.ncbi.nlm.nih.gov/pubmed/22432515
- Persky AM, Brazeau GA, Hochhaus G. Pharmacokinetics of the dietary supplement creatine. Clin Pharmacokinet. 2003;42:557–574. doi: 10.2165/00003088-200342060-00005. https://www.ncbi.nlm.nih.gov/pubmed/12793840
- Spillane M, Schoch R, Cooke M, Harvey T, Greenwood M, Kreider R, Willoughby DS. The effects of creatine ethyl ester supplementation combined with heavy resistance training on body composition, muscle performance, and serum and muscle creatine levels. J Int Soc Sports Nutr. 2009;6:6. doi: 10.1186/1550-2783-6-6. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2649889/
- Knop K, Hoogenboom R, Fischer D, Schubert US. Poly(ethylene glycol) in drug delivery: pros and cons as well as potential alternatives. Angew Chem Int Ed Engl. 2010;49:6288–6308. doi: 10.1002/anie.200902672. https://www.ncbi.nlm.nih.gov/pubmed/20648499
- Camic CL, Hendrix CR, Housh TJ, Zuniga JM, Mielke M, Johnson GO, Schmidt RJ, Housh DJ. The effects of polyethylene glycosylated creatine supplementation on muscular strength and power. J Strength Cond Res. 2010;24:3343–3351. doi: 10.1519/JSC.0b013e3181fc5c5c. https://www.ncbi.nlm.nih.gov/pubmed/21068676
- Herda TJ, Beck TW, Ryan ED, Smith AE, Walter AA, Hartman MJ, Stout JR, Cramer JT. Effects of creatine monohydrate and polyethylene glycosylated creatine supplementation on muscular strength, endurance, and power output. J Strength Cond Res. 2009;23:818–826. doi: 10.1519/JSC.0b013e3181a2ed11. https://www.ncbi.nlm.nih.gov/pubmed/19387397
- Steenge GR, Lambourne J, Casey A, Macdonald IA, Greenhaff PL. Stimulatory effect of insulin on creatine accumulation in human skeletal muscle. Am J Physiol. 1998;275:E974–E979. https://www.ncbi.nlm.nih.gov/pubmed/9843739
- Steenge G, Simpson E, Greenhaff P. Protein- and carbohydrate-induced augmentation of whole body creatine retention in humans. J Appl Physiol. 2000;89:1165–1171. https://www.ncbi.nlm.nih.gov/pubmed/10956365
- Olsen S, Aagaard P, Kadi F, Tufekovic G, Verney J, Olesen JL, Suetta C, Kjaer M. Creatine supplementation augments the increase in satellite cell and myonuclei number in human skeletal muscle induced by strength training. J Physiol. 2006;573:525–534. doi: 10.1113/jphysiol.2006.107359. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1779717/
- Walsh AL, Gonzalez AM, Ratamess NA, Kang J, Hoffman JR. Improved time to exhaustion following ingestion of the energy drink Amino Impact. J Int Soc Sports Nutr. 2010;7:14. doi: 10.1186/1550-2783-7-14. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2861014/
- Jackman SR, Witard OC, Philp A, Wallis GA, Baar K, Tipton KD. Branched-Chain Amino Acid Ingestion Stimulates Muscle Myofibrillar Protein Synthesis following Resistance Exercise in Humans. Frontiers in Physiology. 2017;8:390. doi:10.3389/fphys.2017.00390. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5461297/
- Effects of flooding amino acids on incorporation of labeled amino acids into human muscle protein. Smith K, Reynolds N, Downie S, Patel A, Rennie MJ. Am J Physiol. 1998 Jul; 275(1 Pt 1):E73-8. https://www.ncbi.nlm.nih.gov/pubmed/9688876/
- Postexercise net protein synthesis in human muscle from orally administered amino acids. Tipton KD, Ferrando AA, Phillips SM, Doyle D Jr, Wolfe RR. Am J Physiol. 1999 Apr; 276(4 Pt 1):E628-34. https://www.ncbi.nlm.nih.gov/pubmed/10198297/
- Nonessential amino acids are not necessary to stimulate net muscle protein synthesis in healthy volunteers. Tipton KD, Gurkin BE, Matin S, Wolfe RR. J Nutr Biochem. 1999 Feb; 10(2):89-95. https://www.ncbi.nlm.nih.gov/pubmed/15539275/
- Signaling pathways and molecular mechanisms through which branched-chain amino acids mediate translational control of protein synthesis. Kimball SR, Jefferson LS. J Nutr. 2006 Jan; 136(1 Suppl):227S-31S. https://www.ncbi.nlm.nih.gov/pubmed/16365087/
- Oral leucine administration stimulates protein synthesis in rat skeletal muscle. Crozier SJ, Kimball SR, Emmert SW, Anthony JC, Jefferson LS. J Nutr. 2005 Mar; 135(3):376-82. https://www.ncbi.nlm.nih.gov/pubmed/15735066/
- Effects of leucine and its metabolite β-hydroxy-β-methylbutyrate on human skeletal muscle protein metabolism. Wilkinson DJ, Hossain T, Hill DS, Phillips BE, Crossland H, Williams J, Loughna P, Churchward-Venne TA, Breen L, Phillips SM, Etheridge T, Rathmacher JA, Smith K, Szewczyk NJ, Atherton PJ. J Physiol. 2013 Jun 1; 591(11):2911-23. https://www.ncbi.nlm.nih.gov/pubmed/23551944/
- Signals mediating skeletal muscle remodeling by resistance exercise: PI3-kinase independent activation of mTORC1. Philp A, Hamilton DL, Baar K. J Appl Physiol (1985). 2011 Feb; 110(2):561-8. https://www.ncbi.nlm.nih.gov/pubmed/21071597/
- Activation of mTORC1 by leucine is potentiated by branched-chain amino acids and even more so by essential amino acids following resistance exercise. Moberg M, Apró W, Ekblom B, van Hall G, Holmberg HC, Blomstrand E. Am J Physiol Cell Physiol. 2016 Jun 1; 310(11):C874-84. https://www.ncbi.nlm.nih.gov/pubmed/27053525/
- Leucine supplementation of a low-protein mixed macronutrient beverage enhances myofibrillar protein synthesis in young men: a double-blind, randomized trial. Churchward-Venne TA, Breen L, Di Donato DM, Hector AJ, Mitchell CJ, Moore DR, Stellingwerff T, Breuille D, Offord EA, Baker SK, Phillips SM. Am J Clin Nutr. 2014 Feb; 99(2):276-86. https://www.ncbi.nlm.nih.gov/pubmed/24284442/
- Absence of leucine in an essential amino acid supplement reduces activation of mTORC1 signalling following resistance exercise in young females. Moberg M, Apró W, Ohlsson I, Pontén M, Villanueva A, Ekblom B, Blomstrand E. Appl Physiol Nutr Metab. 2014 Feb; 39(2):183-94. https://www.ncbi.nlm.nih.gov/pubmed/24476474/
- The response of muscle protein synthesis following whole-body resistance exercise is greater following 40 g than 20 g of ingested whey protein. Macnaughton LS, Wardle SL, Witard OC, McGlory C, Hamilton DL, Jeromson S, Lawrence CE, Wallis GA, Tipton KD. Physiol Rep. 2016 Aug; 4(15): https://www.ncbi.nlm.nih.gov/pubmed/27511985/
- Fish oil supplementation suppresses resistance exercise and feeding-induced increases in anabolic signaling without affecting myofibrillar protein synthesis in young men. McGlory C, Wardle SL, Macnaughton LS, Witard OC, Scott F, Dick J, Bell JG, Phillips SM, Galloway SD, Hamilton DL, Tipton KD. Physiol Rep. 2016 Mar; 4(6): https://www.ncbi.nlm.nih.gov/pubmed/27009278/
- Shimomura Y, Yamamoto Y, Bajotto G, Sato J, Murakami T, Shimomura N, Kobayashi H, Mawatari K. Nutraceutical effects of branched-chain amino acids on skeletal muscle. J Nutr. 2006;136:529S–532S. https://www.ncbi.nlm.nih.gov/pubmed/16424141
- Garlick PJ, Grant I. Amino acid infusion increases the sensitivity of muscle protein synthesis in vivo to insulin. Effect of branched-chain amino acids. Biochem J. 1988;254:579–584. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1135117/
- Balage M, Dardevet D. Long-term effects of leucine supplementation on body composition. Curr Opin Clin Nutr Metab Care. 2010;13:265–270. doi: 10.1097/MCO.0b013e328336f6b8. https://www.ncbi.nlm.nih.gov/pubmed/20110810
- Pencharz PB, Elango R, Ball RO. Determination of the tolerable upper intake level of leucine in adult men. J Nutr. 2012;142:2220S–2224S. doi: 10.3945/jn.112.160259. https://www.ncbi.nlm.nih.gov/pubmed/23077191
- Louard RJ, Barrett EJ, Gelfand RA. Effect of infused branched-chain amino acids on muscle and whole-body amino acid metabolism in man. Clin Sci. 1990;79:457–466. https://www.ncbi.nlm.nih.gov/pubmed/2174312
- Borsheim E, Tipton KD, Wolf SE, Wolfe RR. Essential amino acids and muscle protein recovery from resistance exercise. Am J Physiol Endocrinol Metab. 2002;283:E648–E657. https://www.ncbi.nlm.nih.gov/pubmed/12217881
- Stoppani J, Scheett T, Pena J, Rudolph C, Charlebois D. Consuming a supplement containing branched-chain amino acids during a resistance-traning program increases lean mass, muscle strength, and fat loss. J Int Soc Sports Nutr. 2009;6:P1. doi: 10.1186/1550-2783-6-S1-P1.
- Wilson GJ, Layman DK, Moulton CJ, Norton LE, Anthony TG, Proud CG, Rupassara SI, Garlick PJ. Leucine or carbohydrate supplementation reduces AMPK and eEF2 phosphorylation and extends postprandial muscle protein synthesis in rats. Am J Physiol Endocrinol Metab. 2011;301:E1236–E1242. doi: 10.1152/ajpendo.00242.2011. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4395871/
- Suryawan A, Hawes JW, Harris RA, Shimomura Y, Jenkins AE, Hutson SM. A molecular model of human branched-chain amino acid metabolism. Am J Clin Nutr. 1998;68:72–81. https://www.ncbi.nlm.nih.gov/pubmed/9665099
- Fahs CA, Heffernan KS, Fernhall B. Hemodynamic and vascular response to resistance exercise with L-arginine. Med Sci Sports Exerc. 2009;41:773–779. doi: 10.1249/MSS.0b013e3181909d9d. https://www.ncbi.nlm.nih.gov/pubmed/19276857
- Tang JE, Lysecki PJ, Manolakos JJ, MacDonald MJ, Tarnopolsky MA, Phillips SM. Bolus arginine supplementation affects neither muscle blood flow nor muscle protein synthesis in young men at rest or after resistance exercise. J Nutr. 2011;141:195–200. doi: 10.3945/jn.110.130138. https://www.ncbi.nlm.nih.gov/pubmed/21191143
- Volpi E, Kobayashi H, Sheffield-Moore M, Mittendorfer B, Wolfe RR. Essential amino acids are primarily responsible for the amino acid stimulation of muscle protein anabolism in healthy elderly adults. Am J Clin Nutr. 2003;78:250–258. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3192452/
- Alvares TS, Meirelles CM, Bhambhani YN, Paschoalin VM, Gomes PS. L-Arginine as a potential ergogenic aid in healthy subjects. Sports Med. 2011;41:233–248. doi: 10.2165/11538590-000000000-00000. https://www.ncbi.nlm.nih.gov/pubmed/21395365
- Greer BK, Jones BT. Acute arginine supplementation fails to improve muscle endurance or affect blood pressure responses to resistance training. J Strength Cond Res. 2011;25:1789–1794. doi: 10.1519/JSC.0b013e3181e07569. https://www.ncbi.nlm.nih.gov/pubmed/21399536
- McConell GK. Effects of L-arginine supplementation on exercise metabolism. Curr Opin Clin Nutr Metab Care. 2007;10:46–51. doi: 10.1097/MCO.0b013e32801162fa. https://www.ncbi.nlm.nih.gov/pubmed/17143054
- Shao A, Hathcock JN. Risk assessment for the amino acids taurine, L-glutamine and L-arginine. Regul Toxicol Pharmacol. 2008;50:376–399. doi: 10.1016/j.yrtph.2008.01.004. https://www.ncbi.nlm.nih.gov/pubmed/18325648
- Perez-Guisado J, Jakeman PM. Citrulline malate enhances athletic anaerobic performance and relieves muscle soreness. J Strength Cond Res. 2010;24:1215–1222. doi: 10.1519/JSC.0b013e3181cb28e0. https://www.ncbi.nlm.nih.gov/pubmed/20386132
- Sureda A, Cordova A, Ferrer MD, Perez G, Tur JA, Pons A. L-citrulline-malate influence over branched chain amino acid utilization during exercise. Eur J Appl Physiol. 2010;110:341–351. doi: 10.1007/s00421-010-1509-4. https://www.ncbi.nlm.nih.gov/pubmed/20499249
- Hickner RC, Tanner CJ, Evans CA, Clark PD, Haddock A, Fortune C, Geddis H, Waugh W, McCammon M. L-citrulline reduces time to exhaustion and insulin response to a graded exercise test. Med Sci Sports Exerc. 2006;38:660–666. doi: 10.1249/01.mss.0000210197.02576.da. https://www.ncbi.nlm.nih.gov/pubmed/16679980
- Gleeson M. Dosing and efficacy of glutamine supplementation in human exercise and sport training. J Nutr. 2008;138:2045S–2049S. https://www.ncbi.nlm.nih.gov/pubmed/18806122
- Antonio J, Sanders MS, Kalman D, Woodgate D, Street C. The effects of high-dose glutamine ingestion on weightlifting performance. J Strength Cond Res. 2002;16:157–160. https://www.ncbi.nlm.nih.gov/pubmed/11834123
- Haub MD, Potteiger JA, Nau KL, Webster MJ, Zebas CJ. Acute L-glutamine ingestion does not improve maximal effort exercise. J Sports Med Phys Fitness. 1998;38:240–244. https://www.ncbi.nlm.nih.gov/pubmed/9830832
- Colker CM, Swain MA, Fabrucini B, Shi Q, Kalman DS. Effects of supplemental protein on body composition and muscular strength in healthy athletic male adults. Curr Ther Res. 2000;61:19–28. doi: 10.1016/S0011-393X(00)88492-1.
- Candow DG, Chilibeck PD, Burke DG, Davison KS, Smith-Palmer T. Effect of glutamine supplementation combined with resistance training in young adults. Eur J Appl Physiol. 2001;86:142–149. doi: 10.1007/s00421-001-0523-y. https://www.ncbi.nlm.nih.gov/pubmed/11822473
- Camilleri M, Madsen K, Spiller R, Van Meerveld BG, Verne GN. Intestinal barrier function in health and gastrointestinal disease. Neurogastroenterol Motil. 2012;24:503–512. doi: 10.1111/j.1365-2982.2012.01921.x. https://www.ncbi.nlm.nih.gov/pubmed/22583600
- Emerging health properties of whey proteins and their clinical implications. Krissansen GW. J Am Coll Nutr. 2007 Dec; 26(6):713S-23S. https://www.ncbi.nlm.nih.gov/pubmed/18187438/
- Invited review: physiological properties of bioactive peptides obtained from whey proteins. Madureira AR, Tavares T, Gomes AM, Pintado ME, Malcata FX. J Dairy Sci. 2010 Feb; 93(2):437-55. https://www.ncbi.nlm.nih.gov/pubmed/20105516/
- Protective effect of whey protein hydrolysates on H₂O₂-induced PC12 cells oxidative stress via a mitochondria-mediated pathway. Jin MM, Zhang L, Yu HX, Meng J, Sun Z, Lu RR. Food Chem. 2013 Nov 15; 141(2):847-52. https://www.ncbi.nlm.nih.gov/pubmed/23790857/
- Whey protein isolate counteracts the effects of a high-fat diet on energy intake and hypothalamic and adipose tissue expression of energy balance-related genes. McAllan L, Keane D, Schellekens H, Roche HM, Korpela R, Cryan JF, Nilaweera KN. Br J Nutr. 2013 Dec 14; 110(11):2114-26. https://www.ncbi.nlm.nih.gov/pubmed/23731955/
- Effect of whey protein hydrolysates with different molecular weight on fatigue induced by swimming exercise in mice. Liu J, Wang X, Zhao Z. J Sci Food Agric. 2014 Jan 15; 94(1):126-30. https://www.ncbi.nlm.nih.gov/pubmed/23653310/
- Biochemical and metabolic mechanisms by which dietary whey protein may combat obesity and Type 2 diabetes. Jakubowicz D, Froy O. J Nutr Biochem. 2013 Jan; 24(1):1-5. https://www.ncbi.nlm.nih.gov/pubmed/22995389/
- The role of leucine in the regulation of protein metabolism. Garlick PJ. J Nutr. 2005 Jun; 135(6 Suppl):1553S-6S. https://www.ncbi.nlm.nih.gov/pubmed/15930468/
- Post-exercise carbohydrate plus whey protein hydrolysates supplementation increases skeletal muscle glycogen level in rats. Morifuji M, Kanda A, Koga J, Kawanaka K, Higuchi M. Amino Acids. 2010 Apr; 38(4):1109-15. https://www.ncbi.nlm.nih.gov/pubmed/19593593/
- Effect of soy- and whey protein-isolate supplemented diet on the redox parameters of trained mice. Elia D, Stadler K, Horváth V, Jakus J. Eur J Nutr. 2006 Aug; 45(5):259-66. https://www.ncbi.nlm.nih.gov/pubmed/16575496/
- Effects of the dietary amount and source of protein, resistance training and anabolic-androgenic steroids on body weight and lipid profile of rats. Aparicio VA, Sánchez C, Ortega FB, Nebot E, Kapravelou G, Porres JM, Aranda P. Nutr Hosp. 2013 Jan-Feb; 28(1):127-36. https://www.ncbi.nlm.nih.gov/pubmed/23808440/
- A dipeptide and an amino acid present in whey protein hydrolysate increase translocation of GLUT-4 to the plasma membrane in Wistar rats. Morato PN, Lollo PC, Moura CS, Batista TM, Carneiro EM, Amaya-Farfan J. Food Chem. 2013 Aug 15; 139(1-4):853-9. https://www.ncbi.nlm.nih.gov/pubmed/23561181/
- Whey proteins are more efficient than casein in the recovery of muscle functional properties following a casting induced muscle atrophy. Martin V, Ratel S, Siracusa J, Le Ruyet P, Savary-Auzeloux I, Combaret L, Guillet C, Dardevet D. PLoS One. 2013; 8(9):e75408. https://www.ncbi.nlm.nih.gov/pubmed/24069411/
- Whey protein supplementation during resistance training augments lean body mass. Volek JS, Volk BM, Gómez AL, Kunces LJ, Kupchak BR, Freidenreich DJ, Aristizabal JC, Saenz C, Dunn-Lewis C, Ballard KD, Quann EE, Kawiecki DL, Flanagan SD, Comstock BA, Fragala MS, Earp JE, Fernandez ML, Bruno RS, Ptolemy AS, Kellogg MD, Maresh CM, Kraemer WJ. J Am Coll Nutr. 2013; 32(2):122-35. https://www.ncbi.nlm.nih.gov/pubmed/24015719/
- Isoleucine and leucine independently regulate mTOR signaling and protein synthesis in MAC-T cells and bovine mammary tissue slices. Appuhamy JA, Knoebel NA, Nayananjalie WA, Escobar J, Hanigan MD. J Nutr. 2012 Mar; 142(3):484-91. https://www.ncbi.nlm.nih.gov/pubmed/22298573/
- Whey protein hydrolysate augments tendon and muscle hypertrophy independent of resistance exercise contraction mode. Farup J, Rahbek SK, Vendelbo MH, Matzon A, Hindhede J, Bejder A, Ringgard S, Vissing K. Scand J Med Sci Sports. 2014 Oct; 24(5):788-98. https://www.ncbi.nlm.nih.gov/pubmed/23647357/
- In vitro and in vivo safety studies of a proprietary whey extract. Dyer AR, Burdock GA, Carabin IG, Haas MC, Boyce J, Alsaker R, Read LC. Food Chem Toxicol. 2008 May; 46(5):1659-65. https://www.ncbi.nlm.nih.gov/pubmed/18295388/
- Acute and repeated dose (28 days) oral safety studies of ALIBIRD in rats. Anadón A, Martínez MA, Ares I, Castellano V, Martínez-Larrañaga MR, Corzo N, Olano A, Montilla A, Recio I, Martínez-Maqueda D, Miralles B, Fornari T, García-Risco MR, Gonzalez M, Reglero G. J Food Prot. 2013 Jul; 76(7):1226-39. https://www.ncbi.nlm.nih.gov/pubmed/23834798/
- CHEN W-C, HUANG W-C, CHIU C-C, CHANG Y-K, HUANG C-C. Whey Protein Improves Exercise Performance and Biochemical Profiles in Trained Mice. Medicine and Science in Sports and Exercise. 2014;46(8):1517-1524. doi:10.1249/MSS.0000000000000272. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4186725/
- Nutritional physiology of whey and whey components. Barth CA, Behnke U. Nahrung. 1997 Feb; 41(1):2-12. https://www.ncbi.nlm.nih.gov/pubmed/9157293/
- Dietary whey protein lessens several risk factors for metabolic diseases: a review. Sousa GT, Lira FS, Rosa JC, de Oliveira EP, Oyama LM, Santos RV, Pimentel GD. Lipids Health Dis. 2012 Jul 10; 11():67. https://www.ncbi.nlm.nih.gov/pubmed/22676328/
- Antimicrobial activity of whey protein isolate edible films with essential oils against food spoilers and foodborne pathogens. Fernández-Pan I, Royo M, Ignacio Maté J. J Food Sci. 2012 Jul; 77(7):M383-90. https://www.ncbi.nlm.nih.gov/pubmed/22671770/
- National Institute on Drug Abuse. What are anabolic steroids ? https://www.drugabuse.gov/publications/research-reports/anabolic-steroid-abuse/what-are-anabolic-steroids
- National Institute on Drug Abuse. What are steroidal supplements ? https://www.drugabuse.gov/publications/research-reports/anabolic-steroid-abuse/what-are-steroidal-supplements
- National Institute on Drug Abuse. How are anabolic steroids abused ? https://www.drugabuse.gov/publications/research-reports/anabolic-steroid-abuse/how-are-anabolic-steroids-abused
- National Institute on Drug Abuse. Why do people abuse anabolic steroids ? https://www.drugabuse.gov/publications/research-reports/anabolic-steroid-abuse/why-do-people-abuse-anabolic-steroids



{kind=link}