Gluconeogenesis
Gluconeogenesis is the generation of glucose from non-sugar carbon substrates like glycerol, lactate, pyruvate, and glucogenic amino acids 1. The vast majority of gluconeogenesis takes place in your liver and to a smaller extent, in the cortex of kidneys. Gluconeogenesis occurs during periods of fasting, starvation, or intense exercise and is highly endergonic (energy intensive). Gluconeogenesis is often associated with ketosis.
Some organs, such as the brain, the eye, and the kidney, contain tissues that utilize glucose as their preferred or sole metabolic fuel source. During a prolonged fast or vigorous exercise, glycogen stores become depleted, and glucose must be synthesized de novo in order to maintain blood glucose levels.
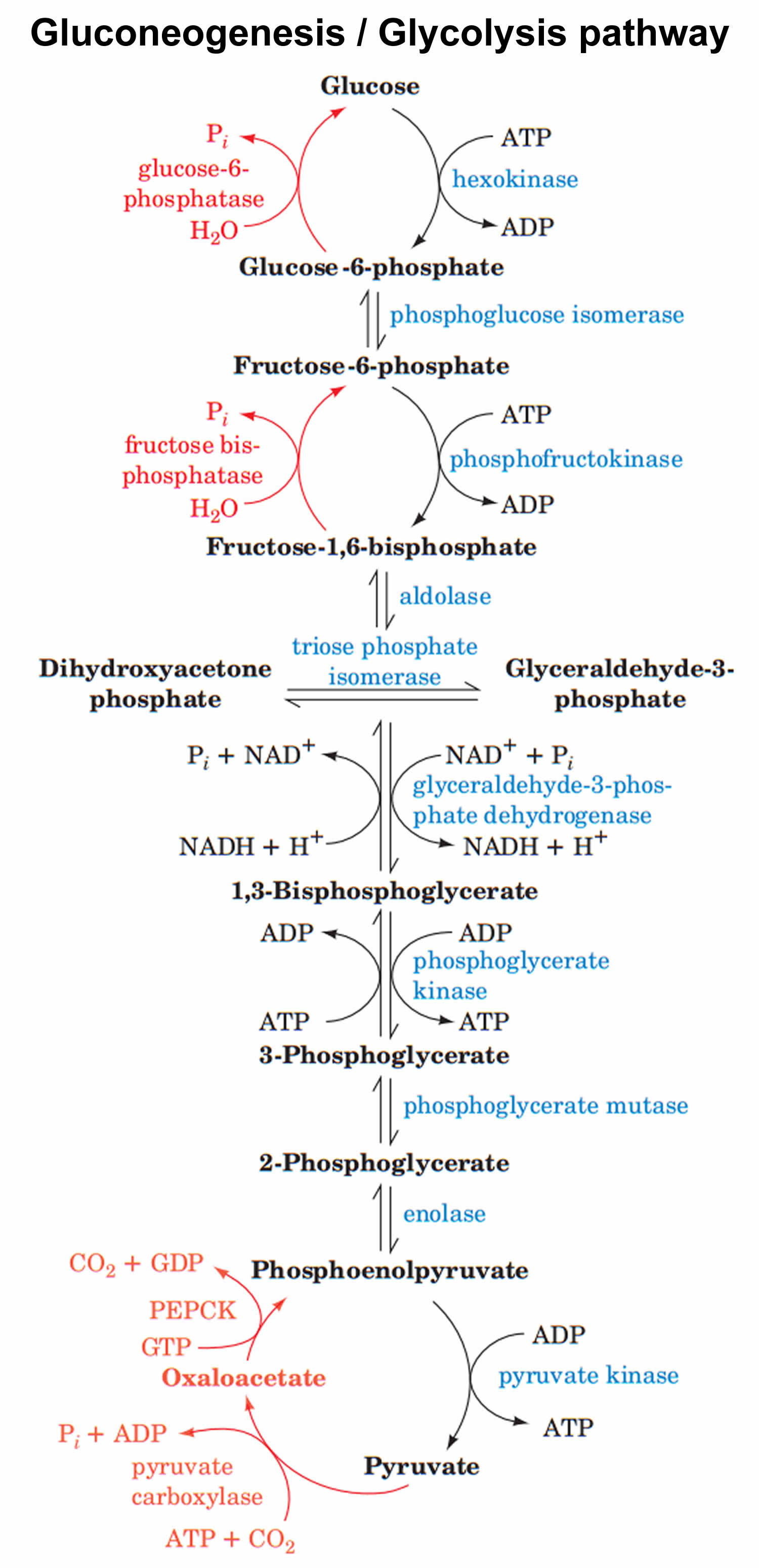
Gluconeogenesis is essentially the reversal of glycolysis 2. However, to bypass the three highly exergonic and essentially irreversible steps of glycolysis, gluconeogenesis utilizes four unique enzymes 1. The enzymes unique to gluconeogenesis are pyruvate carboxylase, phosphoenolpyruvate, carboxykinase, fructose 1,6-bisphosphatase, and glucose 6-phosphatase. Because these enzymes are not present in all cell types, gluconeogenesis can only occur in specific tissues. In humans, gluconeogenesis takes place primarily in the liver and, to a lesser extent, the renal cortex 3.
Several tissues, including the brain, red blood cells, renal medulla, the lens and cornea of the eye, testes, skeletal muscles during exercise, require continuous glucose supply. Among these tissues, the brain uses glucose exclusively in both fed state and fasting state except for prolonged fasting, where the brain uses ketones. Notably, the daily amount of glucose used by the brain accounts for 70% of the total glucose produced by the liver in a normal fasting person 4.
Initially, during the first hours of fasting, hepatic glycogenolysis is the primary source of glucose 5. Glucose stored as glycogen can cover the energy needs roughly for one day; the amount of glucose supplied by glycogen reserves is 190 g while the daily needs for glucose are 160 g 5. After several hours of starvation, gluconeogenesis and glycogenolysis contribute equally to blood glucose. The amount of glucose supplied by glycogen decreases rapidly while the increase in the glucose fraction contributed by gluconeogenesis results in keeping constant the total amount of glucose produced. Estimates are that 54% of glucose comes from gluconeogenesis after 14 hours of starvation, and this contribution raises to 64% after 22 hours and up to 84% after 42 hours 6. However, hours later that glycogen stores deplete, the body uses as glucose sources lactate, glycerol, glucogenic amino acids, and odd chain fatty acids. In prolonged fasting, kidney participation in gluconeogenesis is increased and is responsible for about 40% of total gluconeogenesis 7.
During the first 18 to 24 hours of a fast, the vast majority of gluconeogenesis occurs in the liver. Glucose produced in the liver is shuttled out in circulation and taken up by muscle cells for use in ATP production (Cahill cycle). Following prolonged periods of starvation, however, the kidneys adapt to generate as much as 20% of total glucose produced. Only the liver and kidney can release free glucose from glucose 6-phosphate; other tissues lack the enzyme glucose 6-phosphatase 1.
It is worth mentioning that in certain conditions, such as ischemic strokes and brain tumor development, astrocytes have increased activity of gluconeogenic enzymes, and they use as substrates lactate, alanine, aspartate, glutamate 8.
Gluconeogenesis steps
- Alanine, produced in skeletal muscles by protein catabolism and subsequent transamination reactions, is shuttled out in blood and taken up by the liver. Inside hepatocytes, alanine undergoes transamination into pyruvate, used for gluconeogenesis. In the mitochondrion, pyruvate is carboxylated to form oxaloacetate via the enzyme pyruvate carboxylase, a ligase, adding a carboxyl group on pyruvate to create oxaloacetate 5. Pyruvate carboxylase consumes one ATP molecule, uses as a cofactor biotin (vitamin B7) and uses a CO2 molecule as a source of carbon. Biotin is bound to a lysine residue of pyruvate carboxylase. After ATP hydrolysis, an intermediate molecule pyruvate carboxylase-biotin-CO2 forms, that carboxylates pyruvate forming oxaloacetate. This reaction, apart from forming an intermediate for gluconeogenesis, provides oxaloacetic acid to citric acid cycle or tricarboxylic acid cycle (anaplerotic reaction) 9. In muscle cells, pyruvate carboxylase is used mainly for anaplerotic (act of replenishing citric acid cycle intermediates) reasons. The enzyme also requires magnesium. Pyruvate carboxylation happens in mitochondria; then via malate shuttle, oxaloacetate is being shuttled into the cytosol to be phosphorylated. Malate can cross the inner mitochondrial membrane while oxaloacetic acid cannot. In cytosol along with the oxidation of oxaloacetic acid into malate, NAD+ gets reduced into NADH. The produced NADH is used in a subsequent step when 1,3 bisphosphoglycerate converts into glyceraldehyde-3 phosphate 10. This reaction is unique to gluconeogenesis and is the first of two steps required to bypass the irreversible reaction catalyzed by the glycolytic enzyme pyruvate kinase.
- In red blood cells and other tissues (lens) that lack mitochondria as well as the exercising muscle tissue that favors anaerobic metabolism, glucose is converted to pyruvate and subsequently to lactate. Lactate is secreted into plasma and picked up by the liver for conversion into glucose (Cori cycle) via a redox reaction catalyzed by lactate dehydrogenase 11.
- In the cytosol, oxaloacetate is decarboxylated and rearranged to form phosphoenolpyruvate (PEP) via the enzyme phosphoenolpyruvate carboxykinase (PEPCK), a lyase. Phosphoenolpyruvate carboxykinase (PEPCK) uses guanosine-5′-triphosphate (GTP) as a phosphate donor to phosphorylate oxaloacetate and form phosphoenolpyruvate and magnesium ion as a cofactor. This reaction is unique to gluconeogenesis and is the second of two steps required to bypass the irreversible reaction catalyzed by the glycolytic enzyme pyruvate kinase. Glucocorticoids induce PEPCK gene expression; cortisol after binding its steroid receptor intracellularly moves inside the cell nucleus and binds with its zinc finger domain, the glucocorticoid response element on DNA 12.
- The rest of the reactions are reversible and common with gluconeogenesis. Enolase, a lyase, cleaves carbon-oxygen bonds and catalyzes the conversion of phosphoenolpyruvate (PEP) into 2-phosphoglycerate. Phosphoglycerate mutase, an isomerase, catalyzes the conversion of 2-phosphoglycerate to 3-phosphoglycerate by transferring a phosphate from carbon-2 to carbon-3. Phosphoglycerate kinase using ATP as a phosphate donor and Mg+2 to stabilize with its positive charge the phosphotransfer reaction converts 3-phosphoglycerate to 1,3- bisphosphoglycerate. Glyceraldehyde 3-phosphate dehydrogenase catalyzes the reduction of 1,3-bisphosphoglycerate to glyceraldehyde 3-phosphate. NADH is oxidized as it donates its electrons for the reaction. As described earlier, glycerol phosphate from triglyceride catabolism is converted eventually into dihydroxyacetone phosphate. Triosephosphate isomerase converts dihydroxyacetone phosphate into glyceraldehyde 3-phosphate. Aldolase A converts glyceraldehyde 3-phosphate into fructose-1,6 bisphosphate 3.
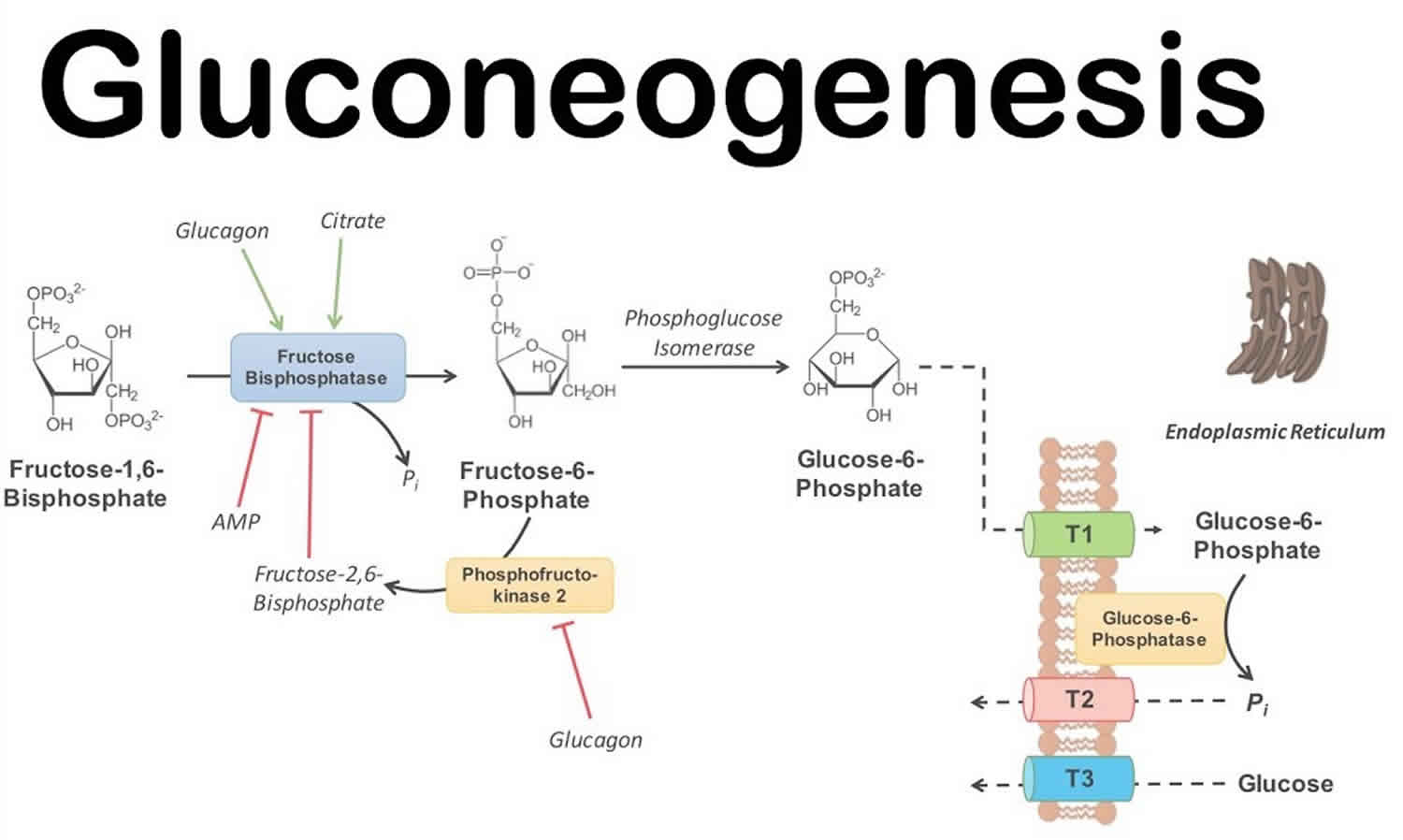
- The following irreversible step involves the conversion of fructose 1,6 bisphosphate into fructose-6 phosphate. This step is important as it is the rate-limiting step of gluconeogenesis. Fructose-1,6 bisphosphatase catalyzes the dephosphorylation of fructose-1,6 bisphosphate, requiring bivalent metal cations (Mg+2, Mn+2); this is a highly regulated step both globally and locally. Locally, increased ATP levels, as well as increased levels of citrate (the first intermediate of tricarboxylic acid cycle), activate the enzyme, while increased AMP and increased fructose-2,6 bisphosphate (F2,6BP) inactivate the enzyme. Glucagon by binding to its receptor, a G protein-coupled receptor (GPCR), activates adenylate cyclase. The resulting increase in cyclic AMP (cAMP) levels leads to the activation of protein kinase A (PKA). PKA phosphorylates fructose 2,6 bisphosphatase (F2,6BPase) and phosphofructokinase-2 (PFK-2). Phosphorylated PFK-2 is inactive while F2,6BPase is active and catalyzes the dephosphorylation of fructose 2,6 bisphosphate. Dephosphorylated F-2,6BP is inactive; hence, it does not have any negative effect on F1,6BPase 13.
- The last irreversible reaction involves glucose-6 phosphatase catalyzing the hydrolysis of glucose-6 phosphate into glucose. This enzyme is expressed primarily in liver as well as in kidneys and intestinal epithelium. The reaction happens in the endoplasmic reticulum of the cells. Muscle cells do not express glucose-6 phosphatase as they produce glucose to maintain their own energy needs 14.
It is worth mentioning that in certain conditions, such as ischemic strokes and brain tumor development, astrocytes have increased activity of gluconeogenic enzymes, and they use as substrates lactate, alanine, aspartate, glutamate.
The major substrates of gluconeogenesis are lactate, glycerol, and glucogenic amino acids 2.
- Lactate is a product of anaerobic glycolysis. When oxygen is limited (such as during vigorous exercise or in low perfusion states) cells must perform anaerobic glycolysis to produce ATP. Cells that lack mitochondria (e.g., erythrocytes) cannot perform oxidative phosphorylation, and as a result rely strictly on anaerobic glycolysis to meet energy demands. Lactate generated from anaerobic glycolysis gets shunted to the liver, where it can be converted back to glucose through gluconeogenesis. Glucose gets released into the bloodstream, where it travels back to erythrocytes and exercising the skeletal muscle to be broken down again by anaerobic glycolysis, forming lactate. This process is called the Cori cycle 3.
- Glycerol comes from adipose tissue. Fatty acids are stored as triglycerides (triacylglycerols) and broken down by the hormone-sensitive lipase in adipose tissue yields free fatty acids and glycerol molecules, the latter of which can circulate freely in the bloodstream until it reaches the liver 15. Glycerol is then phosphorylated by the hepatic enzyme glycerol kinase to yield glycerol phosphate. Next, the enzyme glycerol phosphate dehydrogenase oxidizes glycerol phosphate to yield dihydroxyacetone phosphate, a glycolytic intermediate – an intermediate of gluconeogenesis/ glycolysis pathway- by glycerol phosphate dehydrogenase. Odd-chain fatty acids, in contrast to the ketogenic even- chain fatty acids, are converted with beta-oxidation into propionyl CoA. The latter converts after several steps into methylmalonyl CoA. Methylmalonyl CoA mutase/B12 catalyzes the conversion of the latter into succinyl-CoA. Succinyl-CoA is an intermediate of TCA cycle that is eventually converted into oxaloacetic acid and enters as such the gluconeogenesis pathway. Even-chain fatty acids and purely ketogenic amino acids (leucine, lysine) that convert to acetyl-CoA cannot enter gluconeogenesis as no pathway can reverse the step catalyzed by pyruvate dehydrogenase 16.
- Glucogenic amino acids enter gluconeogenesis via the citric acid cycle. Alanine, produced in skeletal muscles by protein catabolism and subsequent transamination reactions, is shuttled out in blood and taken up by the liver. Inside hepatocytes, alanine undergoes transamination into pyruvate, used for gluconeogenesis. Other gluconeogenic amino acids (e.g., methionine, histidine, valine) as well as gluconeogenic and ketogenic (e.g., phenylalanine, isoleucine, threonine, tryptophane) become transaminated into different intermediates of the gluconeogenic pathway 17. Glucogenic amino acids are catabolized into citric acid cycle metabolites such as alpha-ketoglutarate, succinyl CoA, and fumarate. Through the citric acid cycle, these alpha-ketoacids converts to oxaloacetate, the substrate for the gluconeogenic enzyme PEP carboxykinase.
Figure 1. Gluconeogenesis pathway
Gluconeogenesis regulation
Due to the highly endergonic nature of gluconeogenesis, its reactions are regulated at a variety of levels. The bulk of regulation occurs through alterations in circulating glucagon levels and availability of gluconeogenic substrates. However, fluctuations in catecholamines, growth hormone, and cortisol levels also play a role 18.
- Glucagon is produced by pancreatic alpha cells in response to falling blood glucose levels. Glucagon regulates glucose production by altering the activity of both glycolytic and gluconeogenic enzymes. In response to glucagon, fructose 1,6-bisphosphatase activity is upregulated while its glycolytic counterpart, phosphofructokinase-1, is suppressed 19. Moreover, glucagon binds to an extracellular G protein-coupled receptor (GPCR) that results in the activation of adenylate cyclase and a subsequent increase in the concentration of cAMP 20. cAMP activates cAMP-dependent protein kinase, which then phosphorylates and inactivates the glycolytic enzyme pyruvate kinase. Pyruvate kinase is the enzyme responsible for converting PEP to pyruvate, one of the irreversible reactions of glycolysis. Lastly, glucagon upregulates expression of the gene encoding PEP-carboxykinase, further increasing PEP concentrations and favoring glucose production 20.
- Insulin is a potent inhibitor of gluconeogenesis 21. During a fast, falling insulin levels have a permissive effect on gluconeogenesis. Moreover, a decrease in insulin allows for the catabolism of fat and protein leading to increased gluconeogenic substrate availability 22.
For gluconeogenesis to occur, the ADP/ATP ratio must be very low, since gluconeogenesis is an energy demanding process requiring high energy molecules to be spent in several steps 5. In between meals, during early fasting, when cell via tricarboxylic acid cycle (citric acid cycle) has generated sufficient ATP levels, the increased ATP levels inhibit several highly regulated citric acid cycle (tricarboxylic acid cycle) enzymes (citrate synthase, isocitrate dehydrogenase, a-ketoglutarate dehydrogenase). Acetyl-CoA is the indicator of cells metabolic activity and functions as a gluconeogenesis regulator at a local level. Acetyl-CoA levels back up and allosterically activate pyruvate carboxylase. In this way, the cell makes sure that gluconeogenesis and citric acid cycle (tricarboxylic acid cycle) will not happen simultaneously.
Gluconeogenesis function
The purpose of gluconeogenesis is to maintain blood glucose levels during a fast. In the human body, some tissues rely almost exclusively on glucose as a metabolic fuel source. The brain, for example, requires approximately 120 g of glucose in 24 hours. While the brain is also capable of utilizing ketone bodies as an alternative fuel source, the testes, renal medulla, and erythrocytes all rely exclusively on glucose breakdown through glycolysis. For these tissues to function correctly, a steady influx of glucose into the bloodstream is essential. Hepatic glycogen stores are depleted following a 24-hour fast, after which time gluconeogenesis functions to synthesize glucose de novo from non-hexose precursors and maintain blood glucose levels 3.
Gluconeogenesis clinical significance
Treating hyperglycemia in diabetes
Diabetes is either the result of impaired insulin production or decreased insulin sensitivity. In addition to stimulating glucose uptake from the bloodstream, insulin is also a potent inhibitor of gluconeogenesis. Without adequate insulin production or the ability to respond to insulin properly, gluconeogenesis occurs at an unusually rapid rate, exacerbating hyperglycemia in the diabetic patient 1.
Metformin, the first-line agent for the management of type 2 diabetes, has been shown to suppress hepatic gluconeogenesis through a variety of mechanisms. Metformin activates AMPK, which in turn inhibits hepatic lipogenesis and increases insulin sensitivity. AMPK activation also leads to increased cAMP breakdown, further inhibiting gluconeogenesis 23.
Metformin also appears to directly inhibit glycerol 3-phosphate dehydrogenase, leading to an increase in NADH levels 1. If concentrations of NADH are high enough, the lactate dehydrogenase reaction will favor the formation of lactate over the formation of pyruvate, and lactate will begin to accumulate. Gluconeogenesis is inhibited without the oxidation of lactate to pyruvate.
At high doses, metformin also inhibits complex I of the electron transport chain, impairing ATP production necessary for highly endergonic processes (like gluconeogenesis) to take place 1.
Hypoglycemia as a result of ethanol consumption
Ethanol cannot be eliminated from the human body without changes. To excrete ethanol, it must first be oxidized to form acetaldehyde by the liver enzyme alcohol dehydrogenase, which utilizes NAD+ as an electron acceptor. Next, acetaldehyde must be further oxidized to form acetate (a molecule readily excreted by the body). This reaction, catalyzed by aldehyde dehydrogenase, also requires NAD+ as an electron acceptor. Thus, the metabolism of ethanol results in a significant accumulation of NADH 24.
If concentrations of NADH are high enough, the lactate dehydrogenase reaction will favor the formation of lactate over the formation of pyruvate, and lactate will begin to accumulate. Without the oxidation of lactate to pyruvate, gluconeogenesis is inhibited. As a consequence, heavy ethanol consumption can lead to both lactic acidosis and hypoglycemia 24.
Hypoglycemia in the preterm infant
Preterm infants are at particularly high risk of developing hypoglycemia. Neonates of low birth weight have limited glycogen and fat stores, but also express gluconeogenic enzymes at sub-optimal levels. As such, preterm infants can deplete their energy stores quickly without mounting a proper counter-regulatory response 22.
Von Gierke Disease- Glycogen storage disease type 1
Liver cells lack glucose-6 phosphatase, the enzyme required to release glucose from liver cells by dephosphorylating them. Von Gierke disease is a condition affecting both glycogenolysis and gluconeogenesis since the missing enzyme is common in both pathways resulting in accumulation of glucose-6 phosphate in liver cells. Symptoms include:
- Hepatomegaly and kidney enlargement due to glycogen accumulation
- Severe fasting hypoglycemia since liver cells cannot release glucose in blood postprandially
- Lactic acidosis since accumulated glucose-6 phosphate blocks gluconeogenesis and consequently lactate uptake
- Hypertriglyceridemia, since increased levels of glucose-6 phosphate favor glycolysis and acetyl-CoA production, leading to increased malonyl-CoA synthesis and subsequent inhibition of carnitine acyltransferase 1 (the rate-limiting mitochondrial enzyme of fatty acid beta-oxidation);
Hyperuricemia is the result of increased uric acid production (glc-6P that via HMP shunt is converted into ribose-5P and purines) and decreased uric acid excretion (uric acid competes with lactate for excretion via the same organic acid transporter in proximal renal tubules) 25. Other symptoms include protruding abdomen (hepatomegaly), truncal obesity and short height 26, muscle wasting as well as a rounded doll’s face 25.
Pyruvate Carboxylase deficiency
Pyruvate carboxylase deficiency is a condition where cells lack pyruvate carboxylase or have an altered enzyme and manifest with lactic acidosis, hyperammonemia, and hypoglycemia. Hyperammonemia is due to pyruvate not being converted into oxaloacetic acid. Oxaloacetic acid gets transaminated into aspartate; reduction in aspartate levels results in the reduced introduction of ammonia into the urea cycle 27.
References- Zhang X, Yang S, Chen J, Su Z. Unraveling the Regulation of Hepatic Gluconeogenesis. Front Endocrinol (Lausanne). 2019;9:802. Published 2019 Jan 24. doi:10.3389/fendo.2018.00802 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6353800
- Melkonian EA, Zafar Gondal A, Schury MP. Physiology, Gluconeogenesis. [Updated 2019 Aug 22]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2019 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK541119
- Chung ST, Chacko SK, Sunehag AL, Haymond MW. Measurements of Gluconeogenesis and Glycogenolysis: A Methodological Review. Diabetes. 2015 Dec;64(12):3996-4010.
- Observations on Cerebral Carbohydrate Metabolism in Man. Ann. Intern. Med. 1965 Feb;62:367-71. DOI: 10.7326/0003-4819-62-2-367
- Chourpiliadis C, Mohiuddin SS. Biochemistry, Gluconeogenesis. [Updated 2019 Jul 14]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2019 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK544346
- Chandramouli V, Ekberg K, Schumann WC, Kalhan SC, Wahren J, Landau BR. Quantifying gluconeogenesis during fasting. Am. J. Physiol. 1997 Dec;273(6):E1209-15.
- Gerich JE, Meyer C, Woerle HJ, Stumvoll M. Renal gluconeogenesis: its importance in human glucose homeostasis. Diabetes Care. 2001 Feb;24(2):382-91.
- Yip J, Geng X, Shen J, Ding Y. Cerebral Gluconeogenesis and Diseases. Front Pharmacol. 2016;7:521.
- Adina-Zada A, Zeczycki TN, Attwood PV. Regulation of the structure and activity of pyruvate carboxylase by acetyl CoA. Arch. Biochem. Biophys. 2012 Mar 15;519(2):118-30.
- Jitrapakdee S, St Maurice M, Rayment I, Cleland WW, Wallace JC, Attwood PV. Structure, mechanism and regulation of pyruvate carboxylase. Biochem. J. 2008 Aug 01;413(3):369-87.
- Draoui N, Feron O. Lactate shuttles at a glance: from physiological paradigms to anti-cancer treatments. Dis Model Mech. 2011 Nov;4(6):727-32.
- Stark R, Guebre-Egziabher F, Zhao X, Feriod C, Dong J, Alves TC, Ioja S, Pongratz RL, Bhanot S, Roden M, Cline GW, Shulman GI, Kibbey RG. A role for mitochondrial phosphoenolpyruvate carboxykinase (PEPCK-M) in the regulation of hepatic gluconeogenesis. J. Biol. Chem. 2014 Mar 14;289(11):7257-63.
- Adams A, Redden C, Menahem S. Characterization of human fructose-1,6-bisphosphatase in control and deficient tissues. J. Inherit. Metab. Dis. 1990;13(6):829-48.
- van den Berghe G. Disorders of gluconeogenesis. J. Inherit. Metab. Dis. 1996;19(4):470-7.
- da Silva IV, Rodrigues JS, Rebelo I, Miranda JPG, Soveral G. Revisiting the metabolic syndrome: the emerging role of aquaglyceroporins. Cell. Mol. Life Sci. 2018 Jun;75(11):1973-1988.
- Ferrannini E, Barrett EJ, Bevilacqua S, DeFronzo RA. Effect of fatty acids on glucose production and utilization in man. J. Clin. Invest. 1983 Nov;72(5):1737-47.
- Felig P. The glucose-alanine cycle. Metab. Clin. Exp. 1973 Feb;22(2):179-207.
- Bankir L, Bouby N, Speth RC, Velho G, Crambert G. Glucagon revisited: Coordinated actions on the liver and kidney. Diabetes Res. Clin. Pract. 2018 Dec;146:119-129.
- Droppelmann CA, Sáez DE, Asenjo JL, Yáñez AJ, García-Rocha M, Concha II, Grez M, Guinovart JJ, Slebe JC. A new level of regulation in gluconeogenesis: metabolic state modulates the intracellular localization of aldolase B and its interaction with liver fructose-1,6-bisphosphatase. Biochem. J. 2015 Dec 01;472(2):225-37.
- Borrebaek B, Bremer J, Davis EJ, Davis-Van Thienen W, Singh B. The effect of glucagon on the carbon flux from palmitate into glucose, lactate and ketone bodies, studied with isolated hepatocytes. Int. J. Biochem. 1984;16(7):841-4.
- Honma K, Kamikubo M, Mochizuki K, Goda T. Insulin-induced inhibition of gluconeogenesis genes, including glutamic pyruvic transaminase 2, is associated with reduced histone acetylation in a human liver cell line. Metab. Clin. Exp. 2017 Jun;71:118-124.
- Sharma A, Davis A, Shekhawat PS. Hypoglycemia in the preterm neonate: etiopathogenesis, diagnosis, management and long-term outcomes. Transl Pediatr. 2017 Oct;6(4):335-348.
- Hundal RS, Krssak M, Dufour S, Laurent D, Lebon V, Chandramouli V, Inzucchi SE, Schumann WC, Petersen KF, Landau BR, Shulman GI. Mechanism by which metformin reduces glucose production in type 2 diabetes. Diabetes. 2000 Dec;49(12):2063-9.
- Tsai WW, Matsumura S, Liu W, Phillips NG, Sonntag T, Hao E, Lee S, Hai T, Montminy M. ATF3 mediates inhibitory effects of ethanol on hepatic gluconeogenesis. Proc. Natl. Acad. Sci. U.S.A. 2015 Mar 03;112(9):2699-704.
- Raza M, Arif F, Giyanwani PR, Azizullah S, Kumari S. Dietary Therapy for Von Gierke’s Disease: A Case Report. Cureus. 2017 Aug 08;9(8):e1548.
- Derks TG, van Rijn M. Lipids in hepatic glycogen storage diseases: pathophysiology, monitoring of dietary management and future directions. J. Inherit. Metab. Dis. 2015 May;38(3):537-43.
- Arnold GL, Griebel ML, Porterfield M, Brewster M. Pyruvate carboxylase deficiency. Report of a case and additional evidence for the “mild” phenotype. Clin Pediatr (Phila). 2001 Sep;40(9):519-21.





{kind=link}