What are neutrophils
Neutrophils also known as neutrophilic granulocytes or polymorphonuclear leukocytes are a type of white blood cells (leukocytes) that normally make up the largest number of circulating white blood cells 1. Neutrophils are considered to be the first line of defense during inflammation and infections. Neutrophils play a crucial role in the immune defense against bacterial and fungal pathogens, and they also participate in the development of the inflammatory reaction 2. Neutrophils move into an area of damaged or infected tissue, where they engulf and destroy bacteria or sometimes fungi. Although neutrophils are mostly viewed as playing a beneficial role to the host, their improper activation may also lead to tissue damage during an autoimmune or exaggerated inflammatory reaction 3.
Neutrophils participate in antimicrobial host defense both as the first line of innate immune defense and as effectors of adaptive immunity. Neutrophils are short-lived cells that usually die while performing their antimicrobial function.
A major wave of discoveries during the 1990s and early 2000s made immunologists begin to appreciate the amazing complexity and sophistication of neutrophil functions 1. It became evident that neutrophils release cytokines and contribute to orchestrating the immune/inflammatory response 4. A highly sophisticated machinery directing neutrophil migration 5 and a surprising complexity of neutrophil granules 6 also begun to emerge. Novel but controversial concepts about how neutrophils may fight microbes, including possible regulation of granule enzyme function by ion fluxes 7 and formation of neutrophil extracellular traps (NETs) 8 were also proposed during that period. Both mechanisms have been subject to intense debate. Nevertheless, all those studies indicated that neutrophils use highly sophisticated and complex mechanisms to perform their role in immune defense and inflammation and led to improved and refined models of neutrophil biology 9 (Figure 1).
Key points
- The neutrophil is one of the body’s main cellular mediators of the destruction of microorganisms, and inevitably damages cells and tissues of the host. Neutrophil-mediated tissue destruction is most often a life-saving process, and the host relies on tissue injury as one of the main sources of information that launches inflammation and immunity.
- Large numbers of immature forms of neutrophils, called neutrophilic band cells, are produced by the bone marrow when the demand is high.
- Neutrophils make important contributions to the recruitment, activation and programming of dendritic cells and macrophages. In turn, the adaptive immune system controls the rate of neutrophil production in the bone marrow.
- Neutrophils have important roles in healing wounds, including sterilization of microorganisms, generation of signals that slow the rate of accumulation of more neutrophils, and instigation of a macrophage-based programme that switches the state of damaged epithelium from pro-inflammatory and nonreplicative, to anti-inflammatory and replicative.
- Neutrophil production is coordinated through cytokine production by adaptive immune cells.
- Neutrophil recruitment to sites of infection involves unique molecular interactions in different tissues.
- Recognition of pathogens by neutrophils involves coordination between a repertoire of cellular receptors.
- Killing of pathogens is achieved through the production of toxic metabolites and the release of nuclear contents.
- Heritable disorders of neutrophils provide key insights into molecular mechanisms of neutrophil function.
- Neutrophils play a central role in coordinating the response of other immune effector cells.
- Pathologic interactions between adaptive immune cells and neutrophils are a major contributor to many autoimmune and inflammatory disease states.
- Neutrophils play both positive and negative roles in cancer progression.
- Neutrophils are short-lived cells that die within a limited time after entering the circulation 10. In the absence of infection or inflammation, they die by a spontaneous apoptosis program 10, likely within 1 day (although some investigators propose this time to be up to 5 days) 11. Inflammatory signals are capable of prolonging the lifespan of the cells by several days, during which they release inflammatory mediators and contribute to the orchestration of the inflammatory response 12.
- Accumulating data showed that neutrophils had a variety of important biological functions in both innate and adaptive immunities, far beyond cytotoxicity against pathogens. Neutrophils can differentially switch phenotypes and display distinct subpopulations under different microenvironments. Neutrophils can produce a large variety of cytokines and chemokines upon stimulation. Furthermore, neutrophils directly interact with dendritic cells (DCs), macrophages, natural killer cells, T cells, and B cells so as to either potentiate or down-modulate both innate and adaptive immunity. In the present review, we summarize the recent progress on the functional plasticity and the regulatory ability on immunity of neutrophils in physiological and pathological situations.
- New pathways of tumor-necrosis factor (TNF) signal transduction in neutrophils are being characterized that open up a new way to think of anti-inflammatory therapy, for example, pathways involving tumor-necrosis factor (TNF)-induced increases in intracellular Ca2+ and the ensuing activation of a non-transmembrane form of adenylyl cyclase, termed soluble adenylyl cyclase.
In addition, neutrophils are also found infiltrating many types of tumors 13. Tumor-associated neutrophils have relevant roles in malignant disease. Indeed neutrophils may be potent antitumor effector cells. However, increasing clinical evidence shows tumor-associated neutrophils correlate with poor prognosis. The tumor microenvironment controls neutrophil recruitment and in turn tumor-associated neutrophils help tumor progression. Hence, tumor-associated neutrophils can be beneficial or detrimental to the host 13.
Neutrophils functions
Neutrophils are terminally differentiated cells that develop in the bone marrow where they are also stored for release into the circulation. During their maturation, neutrophils develop several intracellular compartments (secretory granules and vesicles) that store proteins critical for the antimicrobial and proinflammatory missions of the cell 6. Once released into the circulation, neutrophils begin to seek signs of infection and inflammation which lead to a series of events culminating in the migration of neutrophils through the vessel wall and inside the tissue to the site of microbial invasion 5 (see Figure 1).
When encountering infectious particles, neutrophils begin a professional antimicrobial killing program. The infectious particle will be phagocytosed 14 and various noxious agents, including reactive oxygen species generated through the activity of the NADPH oxidase 15, and the contents of intracellular granules 6 are then released into the phagosome, resulting in killing and digestion of the microorganisms (see Figure 1). In cell biology, phagocytosis is the process by which a cell – for example a neutrophil – engulfs a solid particle to form an internal compartment known as a phagosome.
Besides the well-established role of phagocytosis, two additional mechanisms contributing to neutrophil-mediated antimicrobial responses have been proposed during the early 2000s, both of them triggering intensive scrutiny and debate from the scientific community. Segal et al. 7 proposed an interesting molecular interplay whereby ion fluxes triggered by the NADPH oxidase would promote the antimicrobial function of granule proteins. Though many components of the proposed mechanism (such as the electrogenicity of the NADPH oxidase [see Figure 1 inset] providing driving force for other ion currents) are generally accepted, it is still debated whether and to what extent ion fluxes that compensate for the activity of the NADPH oxidase contribute to the antibacterial effects of granule enzymes 16. Another interesting and similarly provocative idea was that, in addition to intraphagosomal killing, neutrophils may also be able to expel their nuclear content (including DNA) complexed with granule proteins into the extracellular space and that this complex, neutrophil extracellular traps (NETs), may inhibit microbial growth without phagocytosis of the pathogen 8. Though neutrophil-derived extracellular DNA has long been known to be present at the site of infection (in part as a component of pus formation) and neutrophil extracellular trap (NET) formation has recently been directly visualized during bacterial infection in vivo 17, the contribution of neutrophil extracellular trap (NET) formation to antibacterial host defense and killing is still under intensive debate 18.
Neutrophils are short-lived cells that die within a limited time after entering the circulation 10. In the absence of infection or inflammation, they die by a spontaneous apoptosis program 10, likely within 1 day (although some investigators propose this time to be up to 5 days) 11. Inflammatory signals are capable of prolonging the lifespan of the cells by several days, during which they release inflammatory mediators and contribute to the orchestration of the inflammatory response 12. However, even under those conditions, neutrophils will soon die by apoptosis (or, possibly, NETosis), contributing to pus formation inside infected tissues. Once neutrophils die, another program for removal of dead neutrophils by macrophages is initiated 19. Parallel to the inflammatory response, biochemical and transcriptional changes leading to the resolution of the inflammation begin 20, and this is further promoted by engulfment of the corpses of dead neutrophils by macrophages 21.
Figure 1. Neutrophils function
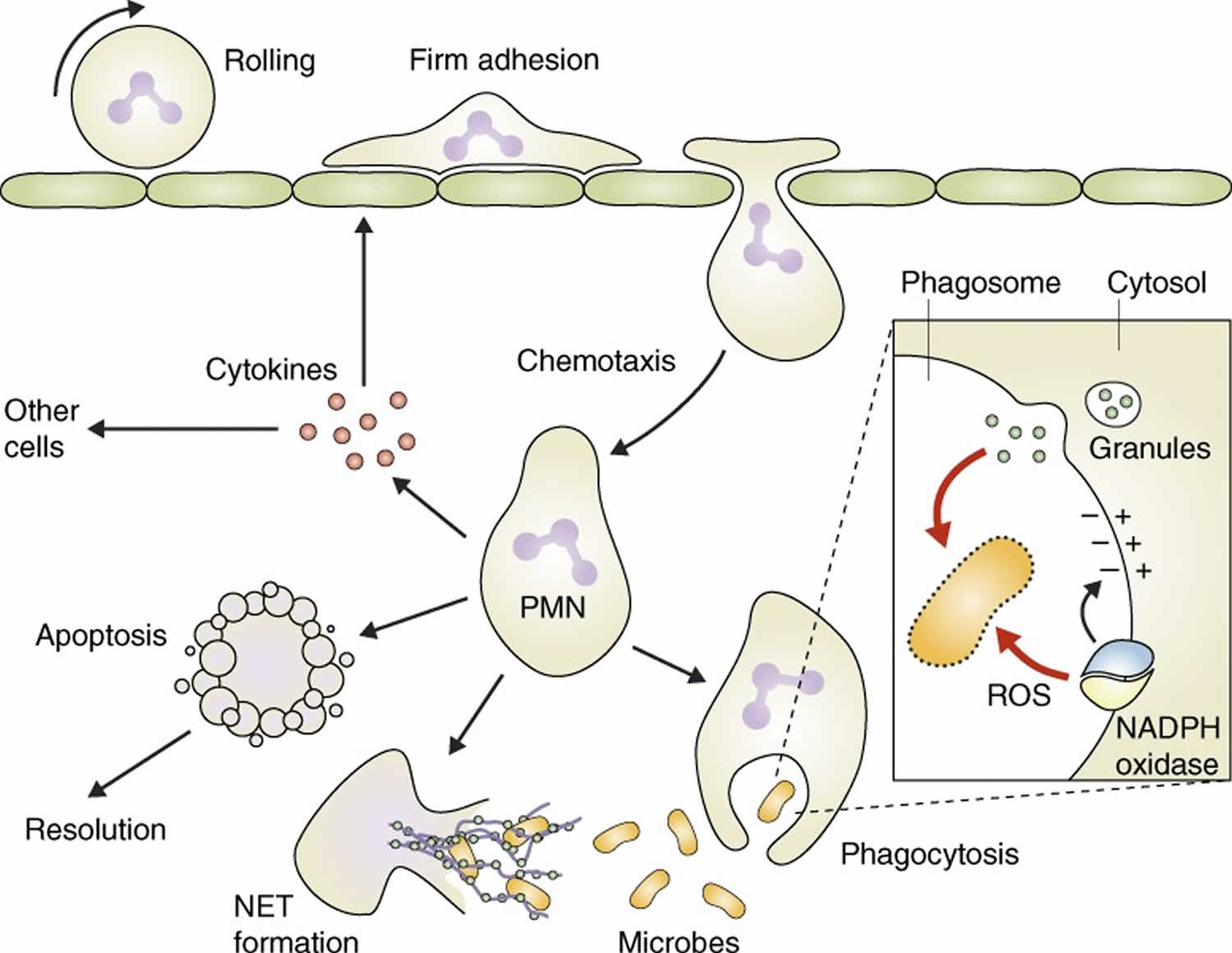
Footnotes: After migrating to the site of inflammation, neutrophils (PMN) phagocytose and digest the invading microbes; release neutrophil extracellular traps (NETs), which likely trap bacteria; and produce cytokines, which contribute to the inflammatory reaction. Once infection is cleared, neutrophils die by apoptosis and trigger an active program to resolve inflammation.
Inset: Cytotoxic functions of neutrophils.
Pathogen killing inside the phagosome occurs by reactive oxygen species (ROS) generated by the NADPH oxidase, as well as by granule enzymes released from intracellular granules. The NADPH oxidase also induces depolarization of the phagosomal membrane, which may be required for providing optimal environment inside the phagosome.
[Source 1]What are white blood cells?
White blood cells (WBCs), also called leukocytes, are cells that circulate in the blood and the lymphatic system that help protect the body against infections. They are an important part of the body’s immune system and also have a role in inflammation, allergic responses, and protection against cancer. A white blood cell differential totals the number of each of the different types of white blood cells in a person’s sample of blood.
There are five types of white blood cells, each with different functions. The differential reveals if the cells are present in normal proportion to one another, if the number of one cell type is increased or decreased, or if abnormal and/or immature cells are present. This information is helpful in diagnosing specific types of illnesses that affect the immune system and the bone marrow.
A differential may be performed in conjunction with a complete blood count (CBC), a test often used as a general health check, or it may be performed in follow-up to abnormal results on a complete blood count (CBC). Most often, a differential is performed on an automated blood analyzer but also may be performed manually by a trained laboratorian who examines a blood smear under a microscope. The values are typically reported as absolute numbers of cells but may be expressed as the relative percentages of the total number of white blood cells.
White blood cells develop from precursor cells produced in the bone marrow.
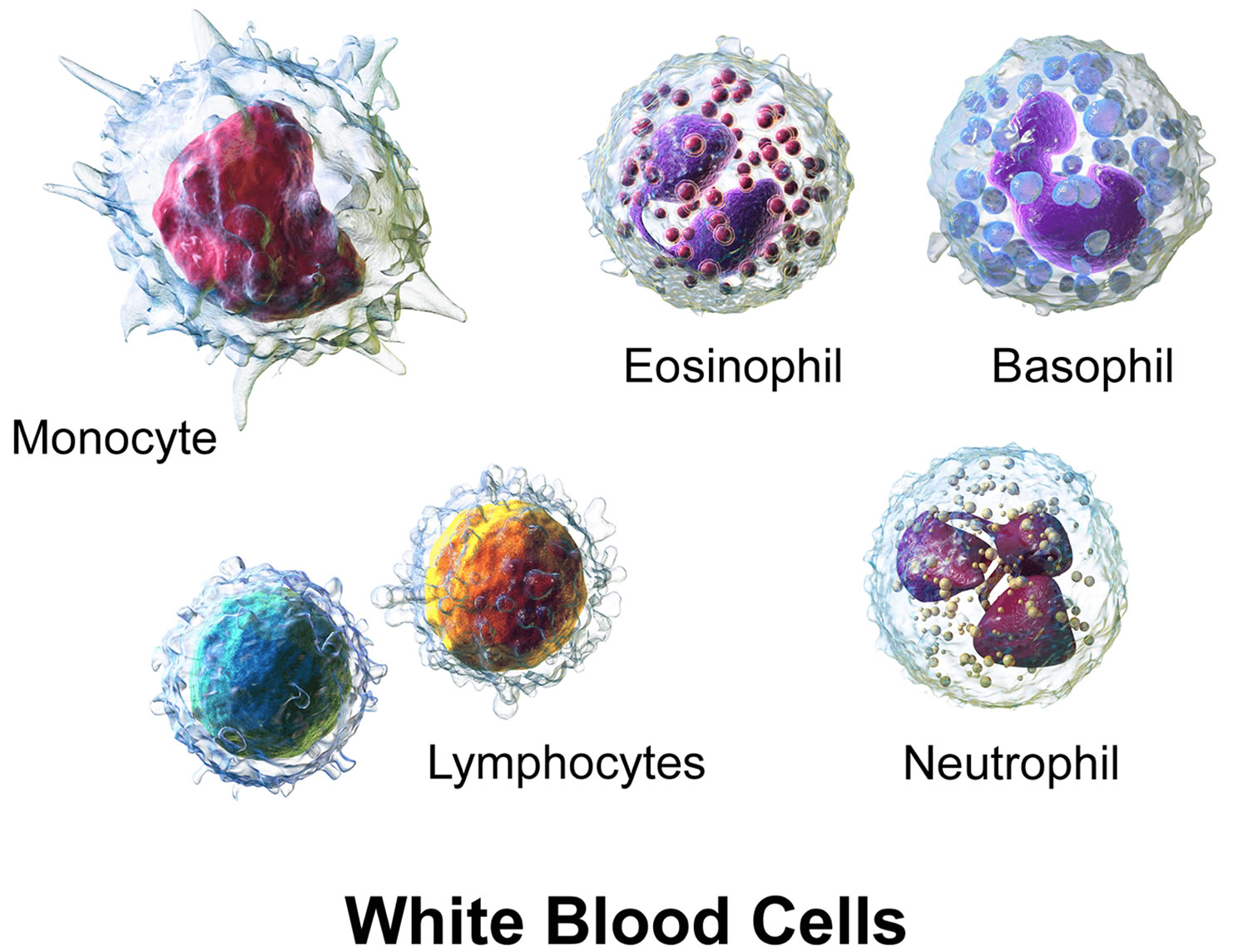
The five different types of white blood cells include:
- Granulocytes—these white blood cells have granules in their cytoplasm. The granules contain chemicals and other substances that are released as part of an immune response. The three types of granulocytes include:
- Neutrophils (neu) normally make up the largest number of circulating white blood cells. They move into an area of damaged or infected tissue, where they engulf and destroy bacteria or sometimes fungi.
- Eosinophils (eos) respond to infections caused by parasites, play a role in allergic reactions (hypersensitivities), and control the extent of immune responses and inflammation.
- Basophils (baso) usually make up the fewest number of circulating white blood cells and are thought to be involved in allergic reactions.
- Lymphocytes (lymphs) exist in both the blood and the lymphatic system. They are divided into three types, but the differential does not distinguish among them. All lymphocytes differentiate from common lymphoid progenitor cells in the bone marrow. The differential counts and reports all lymphocytes together. Separate specialized testing (like immunophenotyping) must be done to differentiate the three types:
- B lymphocytes (B cells) are antibody-producing cells that are essential for acquired, antigen-specific immune responses. Plasma cells are fully differentiated B-cells that produce antibodies, immune proteins that target and destroy bacteria, viruses and other “non-self” foreign antigens.
- T lymphocytes (T cells) finish maturing in the thymus and consist of a few different types. Some T cells help the body distinguish between “self” and “non-self” antigens. Others initiate and control the extent of an immune response, boosting it as needed and then slowing it as the condition resolves. Other types of T cells directly attack and neutralize virus-infected or cancerous cells.
- Natural killer cells (NK cells) directly attack and kill abnormal cells such as cancer cells or those infected with a virus.
- Monocytes (mono), similar to neutrophils, move to an area of infection and engulf and destroy bacteria. They are associated more often with chronic rather than acute infections. They are also involved in tissue repair and other functions involving the immune system.
When there is an infection or an inflammatory process somewhere in the body, the bone marrow produces more white blood cells, releasing them into the blood. Depending on the cause of infection or inflammation, one particular type of white blood cell may be increased as opposed to other types. As the condition resolves, the production of that type of white blood cell subsides and the number drops to normal levels again.
In addition to infections and inflammation, there are a variety of conditions that can affect the production of white blood cells by the bone marrow or their survival in the blood, resulting in either increased or decreased numbers. The differential, along with the other components of the CBC, alerts the healthcare provider to possible health issues. Results are often interpreted in conjunction with additional tests such as a blood smear review, which can reveal the presence of abnormal and/or immature populations of white blood cells.
In a few serious diseases, some immature forms of the cells are released from the bone marrow into the circulation and may be detected by the white blood cell differential. This may occur with bacterial infection, leukemia, bone marrow involvement by solid tumor, myelodysplastic syndrome, or myeloproliferative neoplasms, for example. Some immature cells that may be detected include metamyelocytes, myelocytes, promyelocytes, and/or blasts.
If results indicate a problem, a wide variety of other tests may be performed in order to help determine the cause. A healthcare provider will typically consider an individual’s signs and symptoms, medical history, and results of a physical examination to decide what other tests may be necessary. For example, as needed, a bone marrow biopsy will be performed to evaluate the bone marrow status.
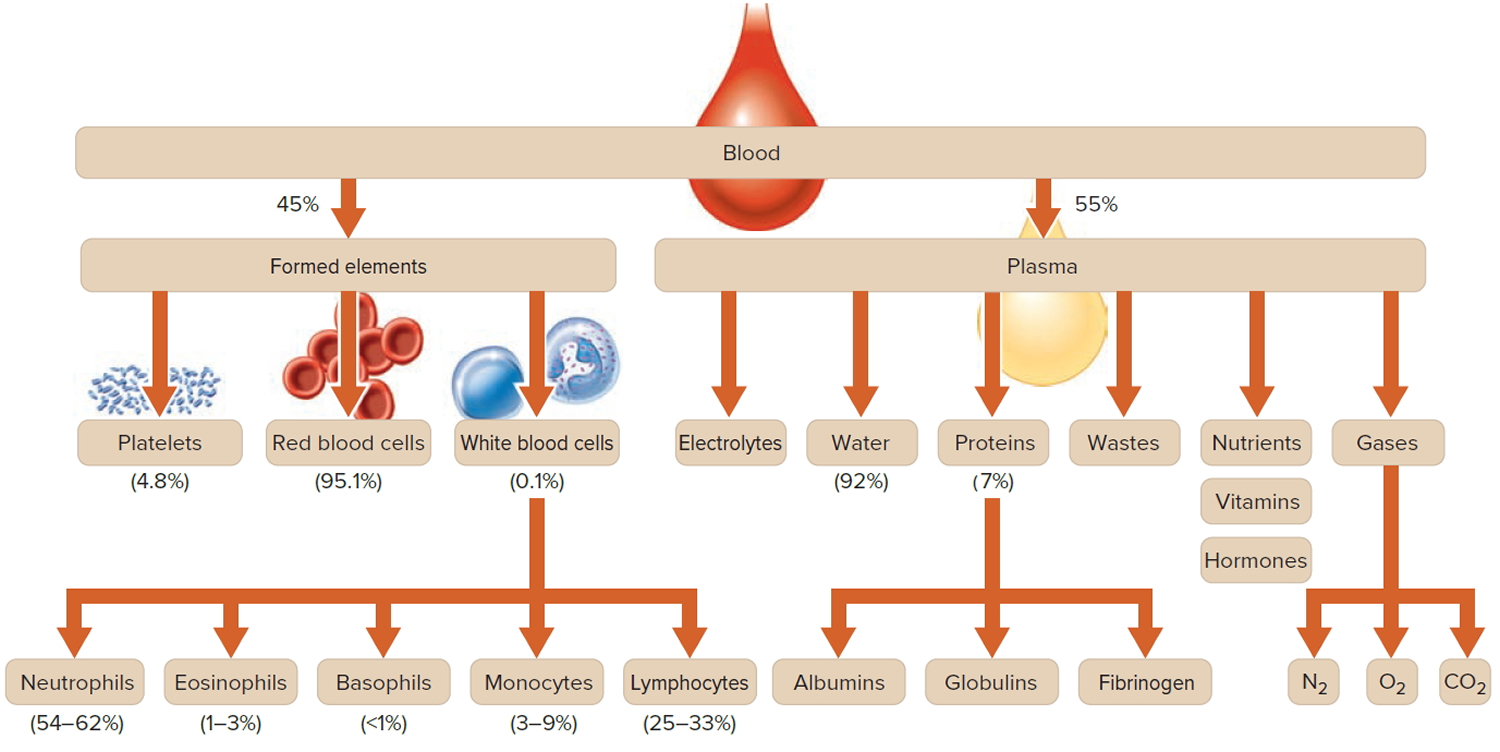
Figure 2. Blood composition
Note: Blood is a complex mixture of formed elements in a liquid extracellular matrix, called blood plasma. Note that water and proteins account for 99% of the blood plasma.
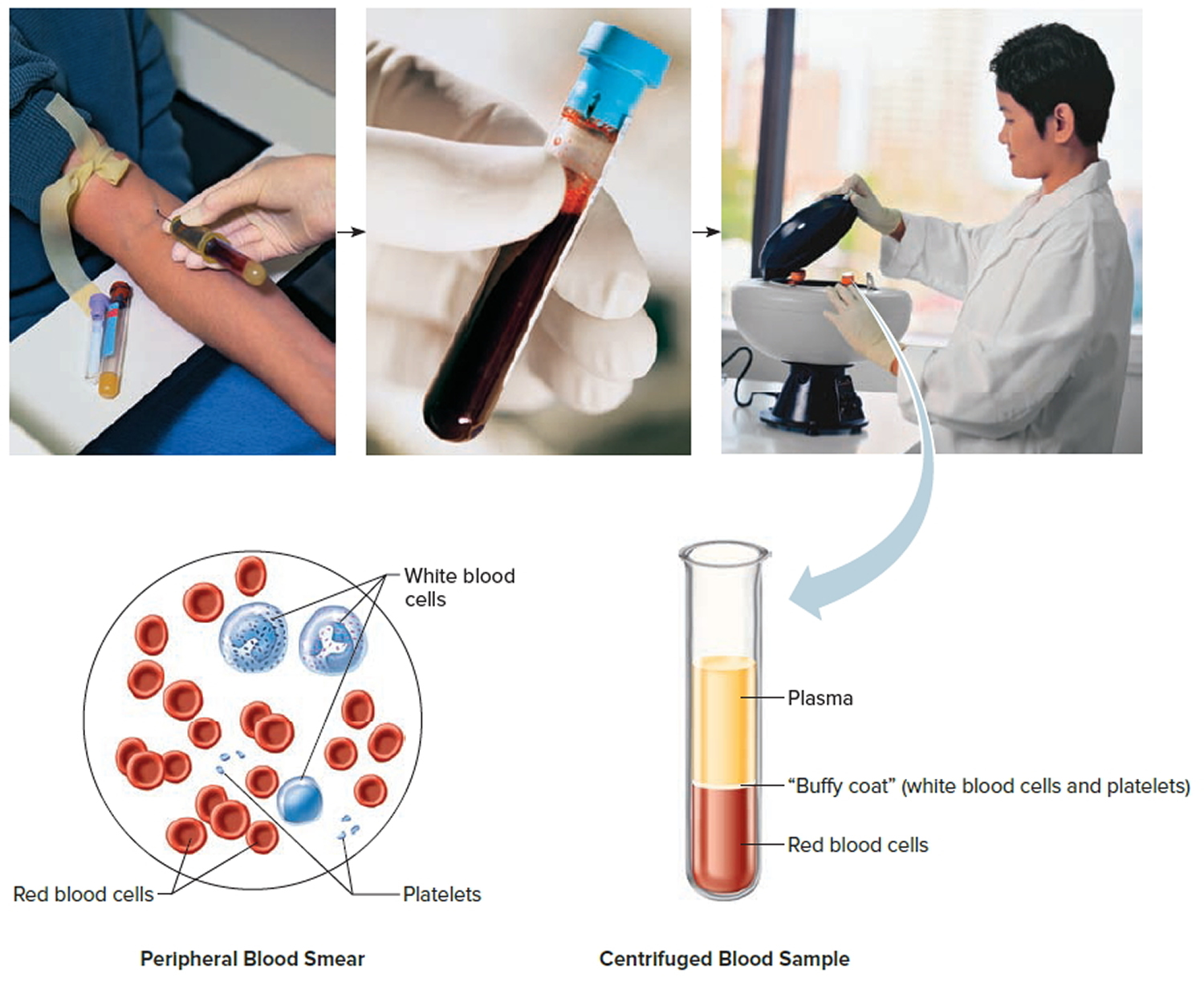
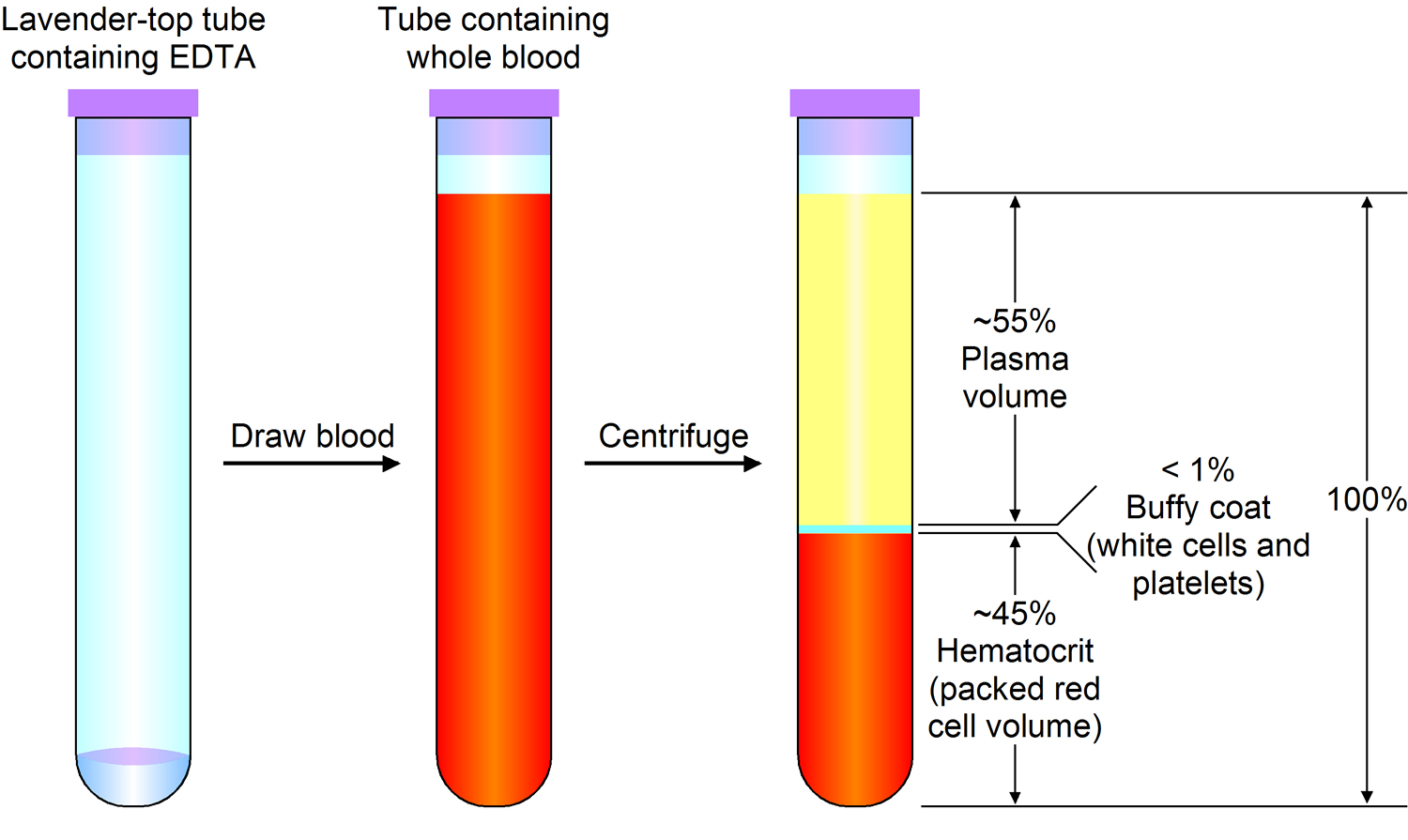
Note: Blood consists of a liquid portion called plasma and a solid portion (the formed elements) that includes red blood cells, white blood cells, and platelets. When blood components are separated by centrifugation, the white blood cells and platelets form a thin layer, called the “buffy coat,” between the plasma and the red blood cells, which accounts for about 1% of the total blood volume. Blood cells and platelets can be seen under a light microscope when a blood sample is smeared onto a glass slide.
Figure 3. Bone marrow anatomy
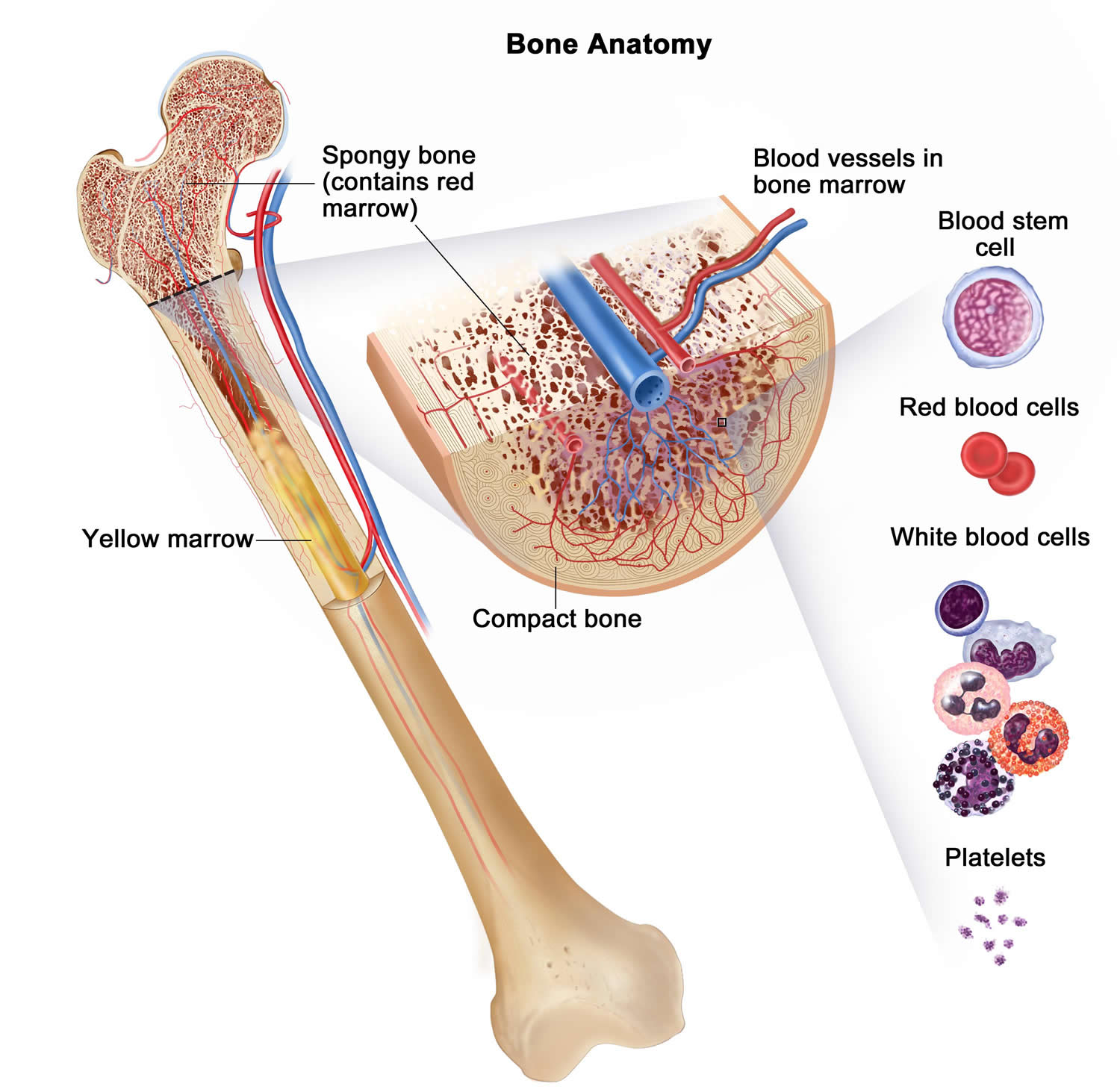
Anatomy of the bone. The bone is made up of compact bone, spongy bone, and bone marrow. Compact bone makes up the outer layer of the bone. Spongy bone is found mostly at the ends of bones and contains red marrow. Bone marrow is found in the center of most bones and has many blood vessels. There are two types of bone marrow: red and yellow. Red marrow contains blood stem cells that can become red blood cells, white blood cells, or platelets. Yellow marrow is made mostly of fat.
Figure 4. White blood cells development. A blood stem cell goes through several steps to become a red blood cell, platelet, or white blood cell
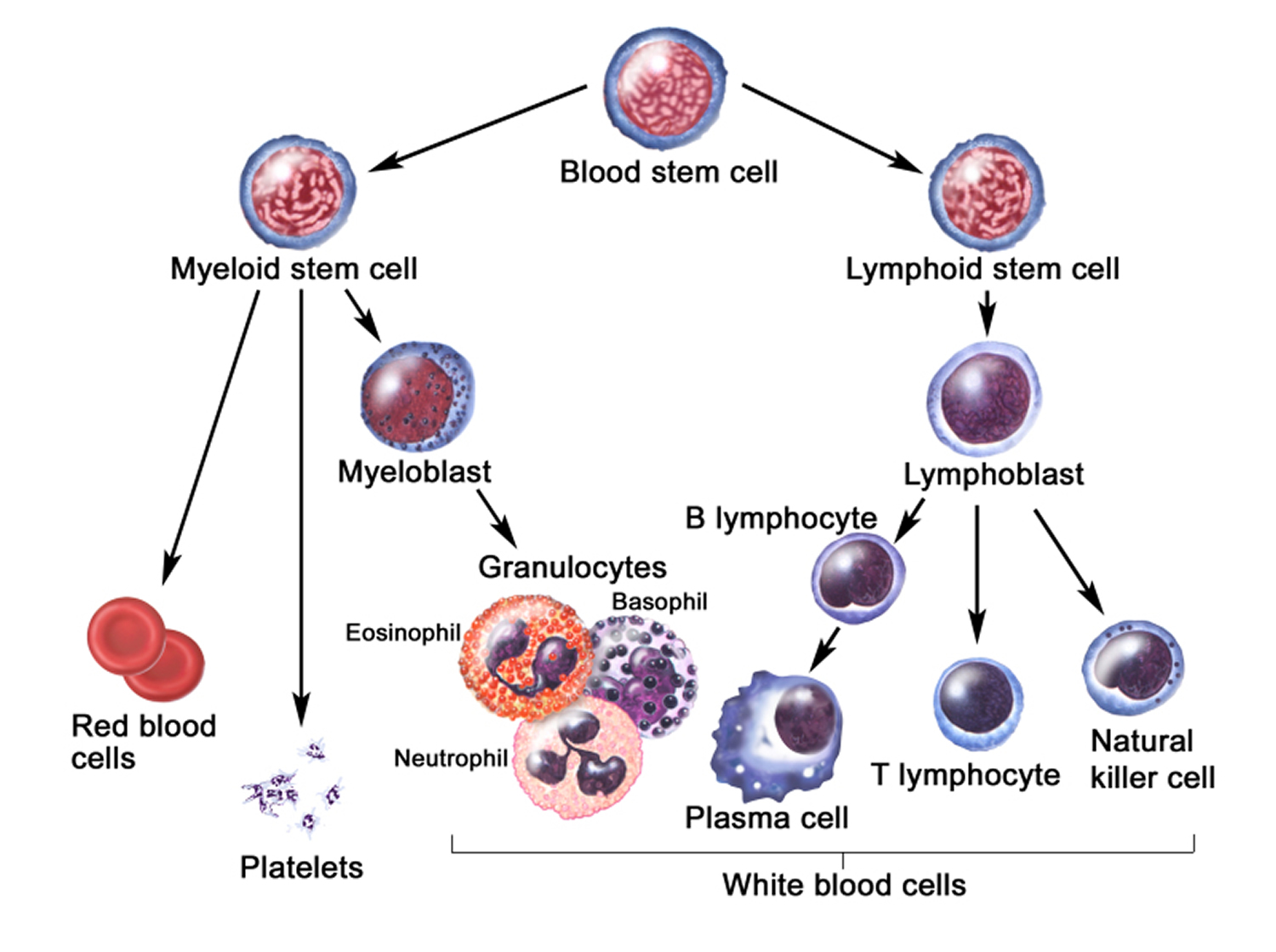
Figure 5. White blood cells development
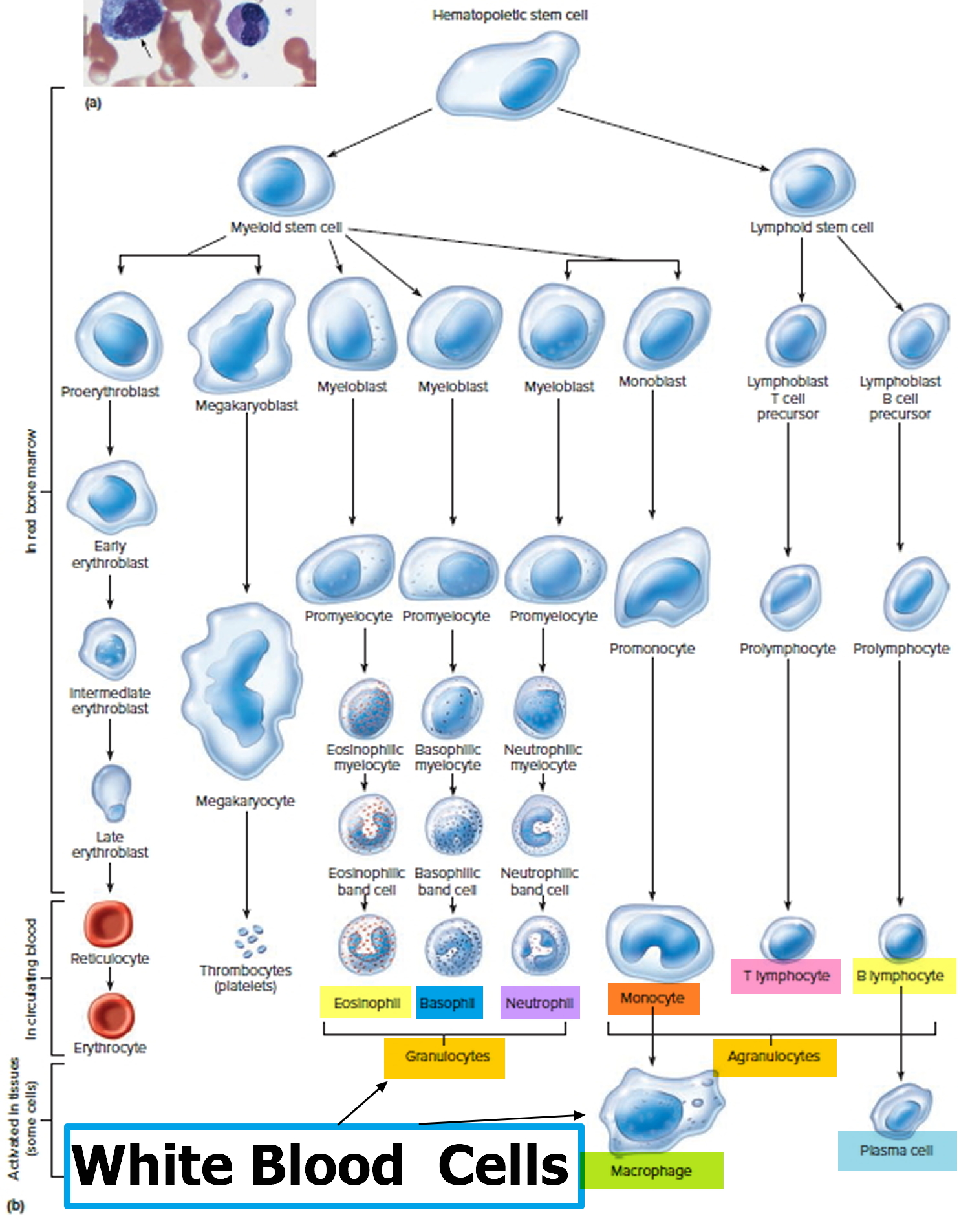
Figure 6. White blood cells
What do neutrophils do
Whereas neutrophils are critical for immunity against bacterial and fungal pathogens, their role in antiviral host defense is much less appreciated. In a recent study, Saitoh et al. 22 proposed a role for neutrophil extracellular traps (NETs) in the control of HIV-1 infection. HIV-1 triggered neutrophil extracellular trap (NET) formation by human neutrophils and neutrophil-derived neutrophil extracellular traps (NETs) were able to capture HIV-1 particles and reduce their efficiency to infect T cells. Most importantly, when neutrophils and T cells were co-cultured, neutrophils were able to reduce the infection efficiency of HIV-1. Pharmacological studies indicated that neutrophil extracellular trap (NET) formation was dependent on TLR7 and TLR8 in endocytic compartments and that the effect of neutrophil-derived neutrophil extracellular traps (NETs) on T cells could be inhibited by DNase treatment, underscoring the role of extracellular DNA. In agreement with those findings, the neutrophil-mediated inhibition of HIV-1 infectivity on T cells required TLR7 and TLR8, and it was suspended by DNase treatment.
Another recent study provided additional evidence for the role of neutrophils and neutrophil extracellular traps (NETs) in antiviral host defense 23. Systemic injection of viral TLR ligands or poxvirus infection led to dramatic accumulation of neutrophils in liver sinusoids that formed aggregates with platelets and released neutrophil extracellular traps (NETs) inside the vessels. Importantly, DNase treatment increased the percentage of liver cells infected with poxviruses, suggesting that extracellular DNA exerts a protective effect against viral infection 23.
Although the aforementioned studies suggest a role for neutrophils and neutrophil extracellular traps (NETs) in antiviral host defense, they also raise several questions. Most studies on NET formation by neutrophils indicate a critical role for NADPH oxidase activity in that process (including the original NET study 8 where NET formation was triggered by phorbol ester-induced activation of neutrophils) and it is difficult to envisage how viral particles would be able to trigger such a strong stimulation of neutrophils. However, because NET formation in the absence of NADPH oxidase activity has recently been shown to occur under milder and more physiological conditions 24, it may be possible that even viral particles trigger NET formation. In addition, Jenne et al. 23 used systemic administration of LPS to induce protection from viral infection. It is unclear whether that response induces specific activation of neutrophils and whether the DNase treatment used only affects neutrophil-derived neutrophil extracellular traps (NETs). Nevertheless, though additional studies in the field are warranted, these two studies (together with additional ones showing priming of virus-specific T cells by neutrophils) open up new avenues on the role of neutrophils in the host defense against viral pathogens 25.
Neutrophils in infection with intracellular pathogens
The traditional view is that neutrophils eradicate extracellular bacteria and fungi, whereas intracellular pathogens such as mycobacteria are primarily dealt with by macrophages. Although prior studies indicated possible roles for neutrophils during mycobacterial infection 26, recent studies provided important novel insights into the role of neutrophils in the fight against intracellular pathogens. Analysis of healthy human individuals who have been in contact with tuberculosis patients revealed strong inverse correlation between the risk of tuberculosis infection and peripheral blood neutrophil counts 27. The authors also showed that ex vivo growth of mycobacteria in whole blood was dramatically increased upon neutrophil depletion, whereas neutrophil-derived antimicrobial peptides inhibited mycobacterial growth from in vitro bacterial cultures 27. Another recent study identified a unique blood transcriptional signature in human tuberculosis patients that closely matched a putative type I and type II interferon-induced gene expression signature in neutrophils 28. This signature was clearly distinguishable from those in other diseases characterized by a major neutrophil component (such as Gram-positive bacterial infections) or by a type I IFN signature (such as SLE). Those two studies suggested that neutrophils actively fight against mycobacteria and may provide protection from infection.
A possible explanation for those findings is that neutrophils fight mycobacteria during and immediately after their entry into the organism. However, this may only be true for the initial contact with mycobacteria when neutrophils are able to prevent infection of the host, while macrophages “take over” when the bacilli reach intracellular compartments 26. Indeed, neutrophils likely play an opposite, pathogenic role during later stages of the disease, through at least two different mechanisms. First, neutrophils may serve as vectors for the systemic dissemination of the pathogens. Mouse neutrophils are able to acquire live mycobacteria at the site of infection and carry those microbes to the draining lymph nodes by migration within the lymphatic vasculature (similar to the migration of dendritic cells in a general immune response) 29. Although this may promote antimicrobial host defense, an alternative scenario is that mycobacteria that escape killing and survive inside neutrophils use them as “Trojan horses” to help disseminate the bacteria to distant organs. This would be similar to the situation in leishmaniasis when neutrophils engulf parasites at the site of infection and deliver them to macrophages, which take up live Leishmania parasites during phagocytosis of apoptotic neutrophils 30. The second mechanism for the contribution of neutrophils to the chronic phase of tuberculosis may be related to their role in infection-induced tissue damage. Cruz et al. 31 suggested that excessive release of IL-17 during repeated mycobacterial exposure leads to neutrophil recruitment and neutrophil-mediated tissue damage in mice. This response appears to be attenuated by IFN-γ production during a normal antimycobacterial immune response 32, indicating that neutrophil accumulation and the resulting tissue damage may be caused by the failure of the immune system to contain the infectious agents.
The dual role of neutrophils in tuberculosis is also reflected in another recent study, suggesting that neutrophils down-regulate inflammation at early stages of mycobacterial infection but contribute to inflammation and mycobacterial clearance during chronic tuberculosis in experimental mice 33. The importance of those findings in human pathology has yet to be assessed, in part because it is uncertain whether IL-10, the cytokine proposed to be responsible for most of the reported findings in mice 33, is also released by human neutrophils 34.
The latest twist in the role of neutrophils in tuberculosis comes from a recent study on zebrafish 35. By monitoring mycobacterial infection, Yang et al. 35 found that neutrophils did not interact with the microbes during the very first phase of infection but engulfed dying infected macrophages during early granuloma formation and killed the surviving intracellular pathogens. That study suggests that efferocytosis (the engulfment of dying cells) not only serves to clean up the site of infection but also contributes to containing the infection, at least in the case of mycobacterial challenge.
Another unexpected role for neutrophils in infectious diseases has been revealed by a recent study on malaria pathogenesis 36. The authors showed that genetic deficiency of IgE- or FcεRI-protected mice from cerebral malaria and death, but not parasitemia after infection with Plasmodium falciparum parasites. Interestingly, FcεRI was dramatically up-regulated on circulating neutrophils after parasitic infection and FcεRI-positive neutrophils became the dominant FcεRI-positive leukocyte population in the brain. Importantly, depletion of neutrophils using the NIMP-R14 anti-Gr1 antibody protected mice both from neurological signs of cerebral malaria and from malaria-induced death, but not from parasitemia, suggesting an important role for neutrophils in malaria pathogenesis 36. Although the mechanism whereby neutrophils contribute to cerebral manifestations of malaria is incompletely understood, the authors suggested that production of proinflammatory cytokines (e.g., IFN-γ) by neutrophils accumulating in the brain of infected animals leads to excessive inflammation and subsequent neurological signs that are characteristic of cerebral malaria.
Collectively, the aforementioned studies suggest that neutrophils may have a much wider (protective or sometimes pathogenic) role during infectious diseases than previously appreciated.
Neutrophils in the regulation of B and T cells
Neutrophils have long been seen as effector cells that have little influence on shaping the adaptive immune response. That view is being challenged by several recent reports.
Marginal zone B cells are unique B cells of the spleen that are located outside the germinal centers and close to the marginal sinus, thus closely positioned to the blood circulation. Their primary function is the rapid initiation of T cell–independent antibody responses to blood-borne microbes. Interestingly, a prior study revealed that neutrophils are one of the major cell types responsible for capturing and transporting circulating bacteria to the splenic marginal zone in mice 37. Additionally, prior human studies showed that activated (but not resting) neutrophils express the B cell–activating cytokine BAFF (also known as BLyS) 38 and that neutrophils are a major source of the BAFF-related, B cell–stimulating cytokine APRIL in B cell lymphomas 39 and mucosal plasma cell niches 40. A recent work by Puga et al. 41 revealed an intricate relationship between neutrophils and marginal zone B cells and proposed that this interaction mediates T cell–independent antibody responses through BAFF and APRIL (Figure 7A). The authors found a large number of neutrophils around the splenic marginal zone in humans, macaques, and mice. In mice, splenic but not circulating neutrophils were able to strongly activate marginal zone B cells, whereas they had limited stimulatory effect on follicular B cells and even inhibited CD4+ T cells. Importantly, splenic neutrophils promoted B cell survival, antibody production, class switching to IgG and IgA, and somatic hypermutation, indicating that they qualify to be bona fide B helper cells. Splenic, but not circulating, neutrophils express large amounts of BAFF, APRIL, and IL-21, and all those cytokines were required for the stimulatory effect on marginal zone B cells. Circulating neutrophils acquired the ability of activating marginal zone B cells upon exposure to splenic sinusoidal endothelial cells in the presence of microbial signals in vitro. The colonization of the splenic marginal zones by neutrophils primarily occurred soon after birth in humans, and marginal zone neutrophils were significantly less abundant in germ-free mice than in normal specific pathogen–free animals, indicating that microbial products, likely originating from the commensal flora, play an important role in the accumulation of splenic marginal zone neutrophils. Importantly, human patients suffering from various congenital defects of neutrophil development or function had normal total circulating B cell numbers but strongly reduced numbers of circulating marginal zone B cells, and their serum levels of antibodies against T cell–independent antigens were significantly reduced despite normal levels of antibodies against T cell–dependent antigens. Collectively, the elegant study of Puga et al. 41 revealed a potentially important immunological and immune-homeostatic mechanism whereby splenic neutrophils may facilitate the antibody response of marginal zone B cells to T cell–independent microbial antigens. This mechanism requires the reprogramming of circulating neutrophils by microbial signals, as well as the neutrophil-derived BAFF, APRIL, and IL-21 cytokines. Further studies will be required to determine the full importance of this mechanism in immune defense and to more closely define the nature and developmental stage of the proposed B cell helper neutrophil population relative to their circulating counterparts.
The role of neutrophils in the regulation of T cell function appears to be more complex and less well understood. Regulation of T cells by myeloid cells primarily occurs via antigen presentation by dendritic cells or macrophages, supported by the migration of antigen-loaded dendritic cells to the draining lymph nodes. Interestingly, human and mouse neutrophils also appear to be able to carry antigens to the lymph nodes by migrating in lymphatic vessels in a CCR7-dependent manner 42. A recent study also showed that vaccinia virus-specific CD8+ T cells emerge in the bone marrow and, based on fluorescence tracking and Ly6G antibody-mediated depletion approaches, identified neutrophils as the cells transporting the antigen from the dermis to the bone marrow 25. Neutrophils also express molecules required for antigen processing and presentation to T cells 43. Late precursors committed to the neutrophil lineage are also important precursors of dendritic cell development. Furthermore, mouse neutrophils are able to cross-prime naive T cells, i.e., present extracellular antigens on MHC class II molecules to CD8+ T cells 43, and also prime antigen-specific Th1 and Th17 cells 44. The transport of antigens from the periphery to the site of T cell priming and the direct priming/activation of antigen-specific T cells in an MHC-restricted manner raise the possibility that neutrophils may possibly even perform some level of classical antigen-presenting cell function (Figure 7C).
Although the aforementioned studies suggest that neutrophils positively regulate antigen-specific T cell responses, several works indicate a negative role for the neutrophil lineage in T cell function. As mentioned above, splenic neutrophils negatively regulate CD4+ T cell function in mice 41. A subset of human neutrophils also inhibits T cell responses during acute systemic inflammatory reactions such as bacterial sepsis 45. Using Ly6G antibody-mediated depletion and G-CSF receptor-deficient mice, a recent study showed that neutrophils prevent the spread of the T cell response to distal lymph nodes during immunization 46. In addition, a large number of immature myeloid cells with inhibitory functions on T cells and other immune function, termed myeloid-derived suppressor cells (MDSCs), can be observed in cancer patients and in various other infectious or inflammatory conditions 47. A subset of those cells (so-called granulocytic MDSCs) share many characteristics with (immature) neutrophils, highlighting the possibility of negative regulation of T cell function by the neutrophil lineage.
Figure 7. Neutrophils interaction with other immune cells
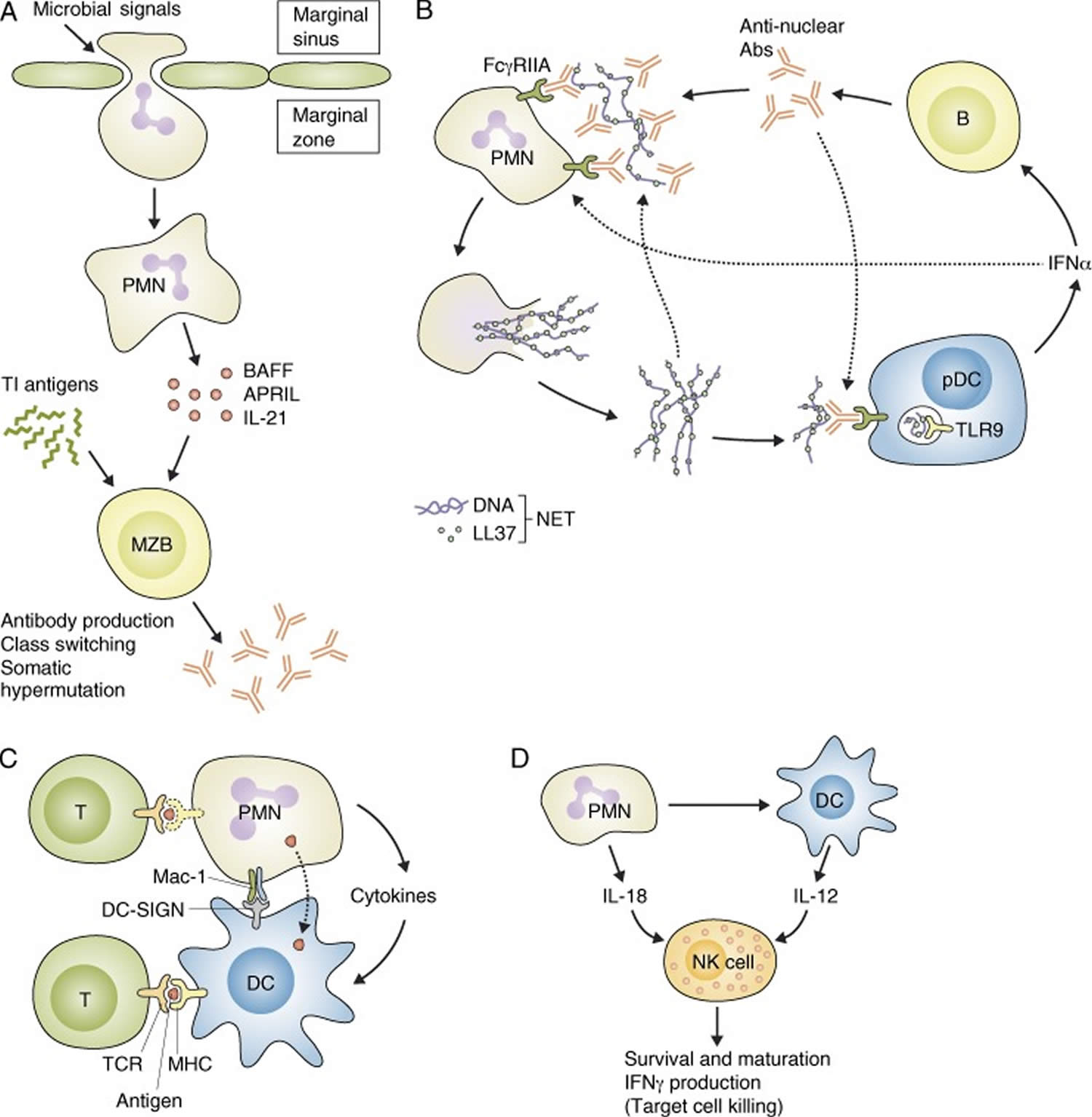
Footnotes: (A) Neutrophils (PMN) accumulate in the splenic marginal zone upon microbial challenge and facilitate the antibody production and maturation of marginal zone B cells (MZB) in response to T cell–independent (TI) antigens. (B) Neutrophils participate in a vicious cycle during autoimmune disease (e.g., SLE) pathogenesis by releasing NETs that trigger pDCs to release IFN-α, which then promotes antibody production by B cells (B). (C) Neutrophils may directly present antigens to T cells (T) and they also activate DCs and deliver antigens to them. (D) Neutrophils and dendritic cells (DCs) jointly activate NK cells (NK).
[Source 1]Regulation of dendritic cells by neutrophils
The effect of neutrophils on adaptive immunity is not only mediated by direct effects on B and T cells, but also by affecting the function of dendritic cells. Indeed, mouse neutrophils are capable of, and required for, the activation of dendritic cells during microbial infection 48. Human and mouse studies suggest that neutrophils directly interact with dendritic cells, which may lead to various responses including dendritic cell maturation and activation, access to antigens captured by neutrophils, or dendritic cell-mediated proliferation and TH1 polarization of T cells 49. The interaction between human neutrophils and dendritic cells has been proposed to be at least in part mediated by a unique interaction between dendritic cell-SIGN and Mac-1 on the neutrophil surface 50 (Figure 7C).
Recent studies have also revealed another unique role of neutrophils in dendritic cell activation during the pathogenesis of autoimmune diseases such as systemic lupus erythematosus (SLE) or type I diabetes (T1D). Characteristic features of SLE, an immune complex–mediated systemic autoimmune disease characterized by autoantibodies against nuclear antigens, include the chronic activation of plasmacytoid dendritic cells (pdendritic cells) leading to constitutive secretion of type I interferons such as IFN-α 51 and the presence of circulating low-density granulocytes reminiscent of immature neutrophils 52. Two recent studies on human patients shed novel light on the role of pdendritic cells, type I interferons, and neutrophils in the core pathogenesis of SLE 53 (Figure 7B). SLE neutrophils were more prone to cell death by NETosis and to release NETs into the extracellular environment. Those NETs contained self-DNA in complex with antimicrobial proteins including LL37 (the C-terminal peptide derived from the human cathelicidin hCAP-18) and were able to activate pdendritic cells through the TLR9 pathway and to trigger type I interferon production. This led to activation of B cells that produced autoantibodies against both self-DNA and the antimicrobial peptides. Type I interferons and immune complexes triggered further activation of neutrophils releasing more NETs 53, establishing a vicious cycle at the core of SLE pathogenesis (Figure 7B). Low-affinity Fcγ-receptors (such as the human FcγRIIA) and TLR7 were required for immune complex-induced neutrophil activation (Garcia-Romo et al., 2011). FcγRIIA may also participate in the uptake of DNA-containing immune complexes by pdendritic cells 54, followed by recognition of DNA-antimicrobial peptide complexes by the TLR9 pathway 55. It should be noted that pdendritic cells do not recognize self-DNA under nonpathological conditions. However, the presence of LL37 in the DNA complexes in NETs prevented degradation of DNA, leading to delivery of DNA to early endosomes of pdendritic cells, where it could activate the TLR9 pathway 53. Therefore, the unique composition of NETs makes them a suitable candidate to trigger pdendritic cell activation in the core of SLE pathogenesis. These results are also in line with a proposed contribution of NET degradation defects to SLE pathogenesis 56, as well as a previous study on the presence of NET formation in autoimmune small vessel vasculitis 57.
The role of neutrophils in pdendritic cell activation and development of autoimmune diseases has also recently been proposed in the pathogenesis of type I diabetes, which is also thought to involve type I interferon production by pdendritic cells 58. Diana et al. 58 reported early infiltration of the pancreas of young nonobese diabetic (NOD) mice with neutrophils, B-1a B cells, and pdendritic cells. Pancreatic β-cell death triggered production and release of anti-DNA IgG antibodies from B-1a cells, which formed DNA–anti-DNA immune complexes and triggered, supposedly in an Fc-receptor–dependent manner, neutrophil NET formation and the release of the murine cathelicidin peptide CRAMP. The DNA–anti-DNA immune complexes and CRAMP then activated pdendritic cells through TLR9 to release IFN-α, which is proposed to trigger further T cell–mediated autoimmune processes. In agreement with the proposed role of neutrophils in the development of type I diabetes, injection of 2-wk-old NOD mice with the NIMP-R14 anti-Gr1 antibody reduced later development of diabetes in those animals 58.
The aforementioned studies suggest an unexpected role for neutrophils in very early phases of autoimmune disease pathogenesis through triggering of type I interferon production by pdendritic cells. There are, however, several open questions that need to be addressed. First, it is widely believed that NET formation requires the NADPH oxidase 59; therefore SLE pathogenesis would be expected to rely on the presence of this enzyme. However, deficiency of the NADPH oxidase component Nox2 exacerbated, rather than inhibited disease pathogenesis in lupus-prone mice 60, arguing against the role of NETs in SLE pathogenesis. This issue is further complicated by NADPH oxidase-independent NET formation proposed by other recent studies 24. Second, although Fc-receptor polymorphisms have been shown to be associated with human SLE 61, no such associations have been reported for type I diabetes (even though NOD mice lacking activating Fc-receptors are partially protected from diabetes development). Therefore, the role of the aforementioned mechanism in diabetes development may not be entirely relevant to human patients. Third, because NETs are likely often formed during bacterial infections 17, it is unclear how autoimmunity is avoided under normal healthy conditions. Nevertheless, the aforementioned mechanism provides an attractive model for the role of neutrophils in autoimmune disease pathogenesis that will merit further investigation. A similar mechanism has also been proposed during development of atherosclerosis, and NETs have also been suggested to participate in the pathogenesis of rheumatoid arthritis by displaying citrullinated autoantigens in an inflammatory environment, further extending the importance of such a pathway in human diseases.
Neutrophils in the regulation of natural killer cell homeostasis
Natural killer (NK) cells are innate lymphocytes with critical roles in the elimination of virus-infected and tumor cells. In vitro interactions between neutrophils and NK cells were described before and a functional cooperation between the two cell types had been proposed 62. The most detailed study tested the role of neutrophils in Legionella pneumophila infection in mice, aiming to understand why neutrophils, which are not capable of killing Legionella species themselves, still play a role in clearing L. pneumophila infection 63. The authors confirmed that depletion of neutrophils by anti-Gr1 antibodies attenuated host defense against Legionella but it also blocked IFN-γ production by NK cells. Using elegant genetic approaches and mixed bone marrow chimeric studies, they showed that IFN-γ production by NK cells was directly stimulated by IL-18 released from neutrophils in response to Legionella infection 63, suggesting that interactions between neutrophils and NK cells may be of functional importance (Figure 7D).
A recent study has provided novel and unexpected genetic evidence for the role of neutrophils in regulating NK cell homeostasis and function 64. The authors set up a random chemical mutagenesis screen to identify novel mutant mouse strains with defective NK cell development and function. They identified a mutation called Genista, which led to hyporesponsive NK cells as indicated by reduced in vitro and in vivo responses to target cells and ligation of NK cell–activating receptors 64. Interestingly, adoptive transfer studies revealed hyporesponsiveness of wild-type NK cells transferred to Genista recipients, indicating that the NK cell phenotype was, at least in part, caused by an environmental, non–cell autonomous defect. In parallel studies, the Genista mutation was identified to be a hypomorphic missense mutation of the Gfi-1 transcription factor, leading to defective terminal differentiation of neutrophils (as indicated by the absence of fully mature neutrophils in the periphery) without negatively affecting T or B cell lineages or other myeloid (monocyte, macrophage, dendritic cell) compartments 64. This was in line with the similar, though more severe, defect in neutrophil maturation in mice with complete disruption of the Gfi-1–encoding gene 65 and congenital neutropenia in human patients carrying a dominant–negative Gfi-1 mutation 66. To further confirm the role of neutrophils in NK cell function, the authors showed that depletion of neutrophils by an anti-Ly6G antibody also reduced the functional responsiveness of NK cells 64. Further studies revealed that both the Genista mutation and the Ly6G antibody-mediated depletion led to defective terminal maturation and poor survival of NK cells. The authors also reported that patients suffering from various forms of congenital or acquired (autoimmune) neutropenia had higher percentages of NK cells showing an immature phenotype and reduced responsiveness to target cells 64. Interestingly, one of the patients with autoimmune neutropenia showed a remission during the course of the study, during which normalization of peripheral neutrophil count and NK cell function occurred in parallel. Collectively, those results suggest that neutrophils likely play a role in regulating the terminal differentiation and functional responsiveness of NK cells (Figure 7D). The colocalization and direct physical interaction of the two cell types in mouse lymphoid organs 64 and inflamed human tissues 67 suggest that neutrophils may directly regulate the NK cell compartment.
Although the aforementioned studies indicate an important role for neutrophils in the regulation of NK cell function, the contribution of a third cell type (such as dendritic cells) in this interaction cannot be excluded. Indeed, besides neutrophil-derived IL-18, IFN-γ production by NK cells upon Legionella infection also required IL-12 derived from dendritic cells as a result of their reciprocal regulation with NK cells 63 (Figure 7D). A tripartite complex formed by neutrophils, NK cells and a minor subset of dendritic cells (slandendritic cells) has also been observed in human samples (including inflammatory lesions in Crohn’s disease and psoriasis) and was proposed to promote IFN-γ production by NK cells 67.
Collectively, neutrophils have recently emerged as potentially important regulators of NK cell homeostasis and activation, possibly in close relationship with dendritic cells.
Neutrophils in allergy and anaphylaxis
Allergic and anaphylactic reactions are adaptive immune responses triggered by repeated exposure to allergens, and are typically characterized by vascular reactions such as vasodilation and edema formation. Several recent studies suggest unexpected roles for neutrophils in those reactions.
The prevailing paradigm of anaphylaxis development is that allergen-induced cross-linking of IgE molecules bound to Fcε-receptors on mast cells and basophils triggers histamine release and subsequent systemic vasodilation 68. Although this mechanism is likely responsible for the majority of anaphylactic reactions, several observations indicate that alternative mechanisms, possibly mediated by IgG and cellular lineages other than mast cells and basophils, may also be at play 69. Jönsson et al. 70 identified neutrophils as important players of active and IgG-induced passive systemic anaphylaxis in experimental mice (Figure 8A). Passive systemic anaphylaxis triggered by IgG antibodies was blocked by neutrophil depletion using the RB6-8C5 anti-Gr1 antibody. Active systemic anaphylaxis was partially reduced by anti-Gr1 antibodies alone, and it was nearly completely blocked by simultaneously depleting both neutrophils and basophils 70. Interestingly, mice lacking all activating Fcγ- and Fcε-receptors except FcγRIV were able to mount normal active anaphylactic reactions which were, however, completely blocked by anti-Gr1 antibodies. Adoptive transfer of wild-type neutrophils rescued anaphylaxis development in FcR γ-chain-deficient animals (which lack all activating Fcγ- and Fcε-receptors including FcγRIV, and fail to mount active systemic anaphylaxis). Those results suggest that neutrophils are responsible for the FcγRIV-mediated component of active systemic anaphylaxis in mice and that this component is sufficient to trigger a full-blown anaphylaxis response. Additional experiments indicated that the neutrophil-dependent anaphylactic reaction was mediated primarily by PAF rather than histamine 70. Importantly, active systemic anaphylaxis could be restored by adoptive transfer of human neutrophils to FcRγ−/− animals or by transgenic expression of human FcγRIIA in FcRγ−/− mice 70,, suggesting that human neutrophils may also be able to mediate systemic anaphylactic reactions.
Figure 8. Neutrophils in vascular diseases
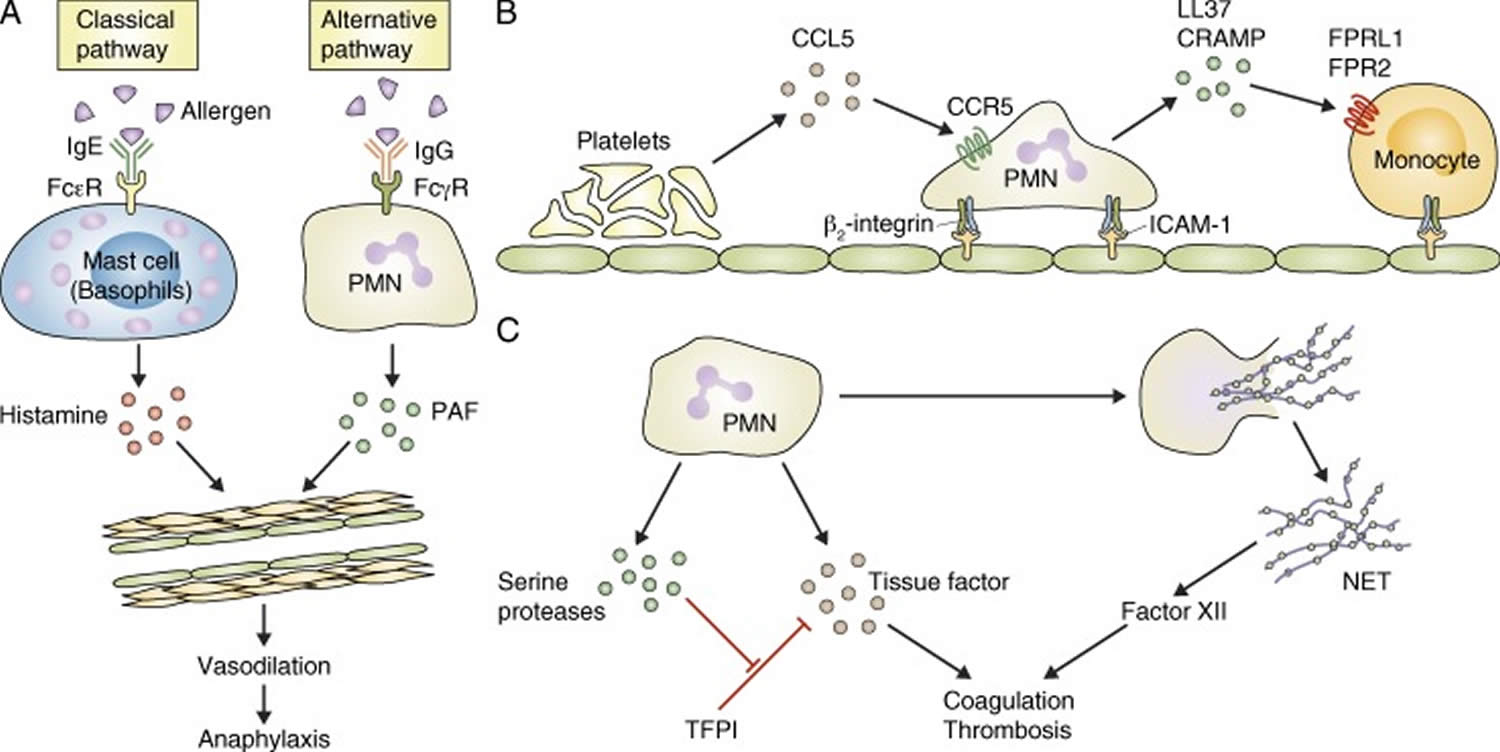
Footnotes: (A) Neutrophils (PMN) mediate an alternative pathway of anaphylactic reaction which is distinct from the classical pathway mediated by mast cells or basophils. (B) Sequential recruitment of platelets, neutrophils, and monocytes (Mono) participates in the pathogenesis of atherosclerosis. (C) Neutrophils contribute to coagulation and thrombosis by releasing tissue factor and inhibitors of the tissue factor pathway inhibitor (TFPI), as well as the procoagulant NETs.
[Source 1]The role of neutrophils in atherosclerosis
Cardiovascular diseases are among the most prevalent human diseases and are the leading causes of death in developed countries. Several recent studies suggest that neutrophils play an important role in those diseases, in particular the development and progression of atherosclerosis and the pathogenesis of thrombosis.
It has long been appreciated that atherosclerosis results from chronic inflammation of the vascular wall 71. Recent reports indicate that neutrophils are present in unique areas of atherosclerotic plaques in both humans and mice 72 and that their presence correlates with poor prognosis and signs of plaque destabilization in human patients 73. Importantly, depletion of neutrophils by antibodies against Ly6G strongly reduced the area of atherosclerotic lesions in apolipoprotein E-deficient (ApoE−/−) mice fed a high-fat diet for 4 wk, suggesting an important role for neutrophils in atherosclerotic plaque formation 74. A recent study provided additional support for the role of neutrophils in atherosclerosis, showing that the genetic deficiency of CRAMP, a member of the cathelicidin family of antimicrobial peptides expressed in neutrophils, leads to reduced atherosclerotic lesion size in ApoE−/− mice fed a high-fat diet 75. CRAMP and the homologous human cathelicidin peptide LL37 (the C-terminal portion of its precursor hCAP-18) promoted the adhesion of classical (inflammatory) monocytes to the vessel wall in mice, and this was inhibited by antagonists of formyl-peptide receptors, the receptors thought to be responsible for cathelicidin-mediated leukocyte migration 75. In addition, activation of pDCs by complexes of cathelicidins with extracellular DNA from neutrophils or other dying cells may also contribute to atherosclerosis development in mice 76 in a manner similar to that proposed for the pathogenesis of SLE and T1D 77. The recruitment of neutrophils to the inflamed mouse arterial wall could also be reduced by platelet depletion or by the genetic deletion of CCR5, the receptor for the platelet-derived CCL5 (RANTES) chemokine 74 which is also required for atherosclerotic plaque formation 78. The release and deposition of CCL5 appears to be dependent on P-selectin on platelets 74. Platelets were previously proposed to participate in atherosclerotic plaque formation 79 and platelet-derived CCL5 was shown to promote monocyte adhesion to the luminal surface of early atherosclerotic lesions in the inflamed arterial wall in mice 79. Mice lacking P-selectin, CD18, or ICAM-1 were previously shown to have reduced atherosclerotic plaque formation 80. Collectively, atherosclerotic plaque formation likely involves the sequential recruitment of platelets, neutrophils, and monocytes, whereby platelet-derived CCL5, acting on CCR5, is at least partially responsible for neutrophil recruitment and neutrophil promotion of monocyte recruitment by cathelicidin-induced activation of formyl-peptide receptors (Figure 8B). Platelet P-selectin is likely involved in platelet deposition (although an additional role of endothelial P-selectin in adhesion of leukocyte rolling cannot be excluded) and neutrophil/monocyte adhesion is likely mediated by interaction between β2-integrins and ICAM-1 (intercellular adhesion molecule 1).
What is neutrophils absolute
A routine blood smear identifies the five types of white blood cells—neutrophils, lymphocytes, monocytes, eosinophils, and basophils. Neutrophils make up about 55% to 70% of the total white blood cell count. Their primary function is phagocytosis—a process by which they engulf and digest microorganisms and cell fragments.
Acute bacterial infection and trauma stimulate neutrophil production, which elevates the white blood cell count. Significant stimulation of neutrophil production causes immature neutrophil forms, called bands or stab cells, to enter the circulation.
Absolute neutrophil count is an essential monitoring parameter for cancer patients at risk for bone marrow suppression caused by chemotherapy, radiation therapy, or bone marrow transplant. If your patient’s absolute neutrophil count is abnormally low, be sure to practice meticulous hand hygiene—the single most important way to prevent infection in these patients.
Normal and abnormal absolute neutrophil count values
A healthy person has an absolute neutrophil count between 2,500 and 6,000/mm³ or per microliter.
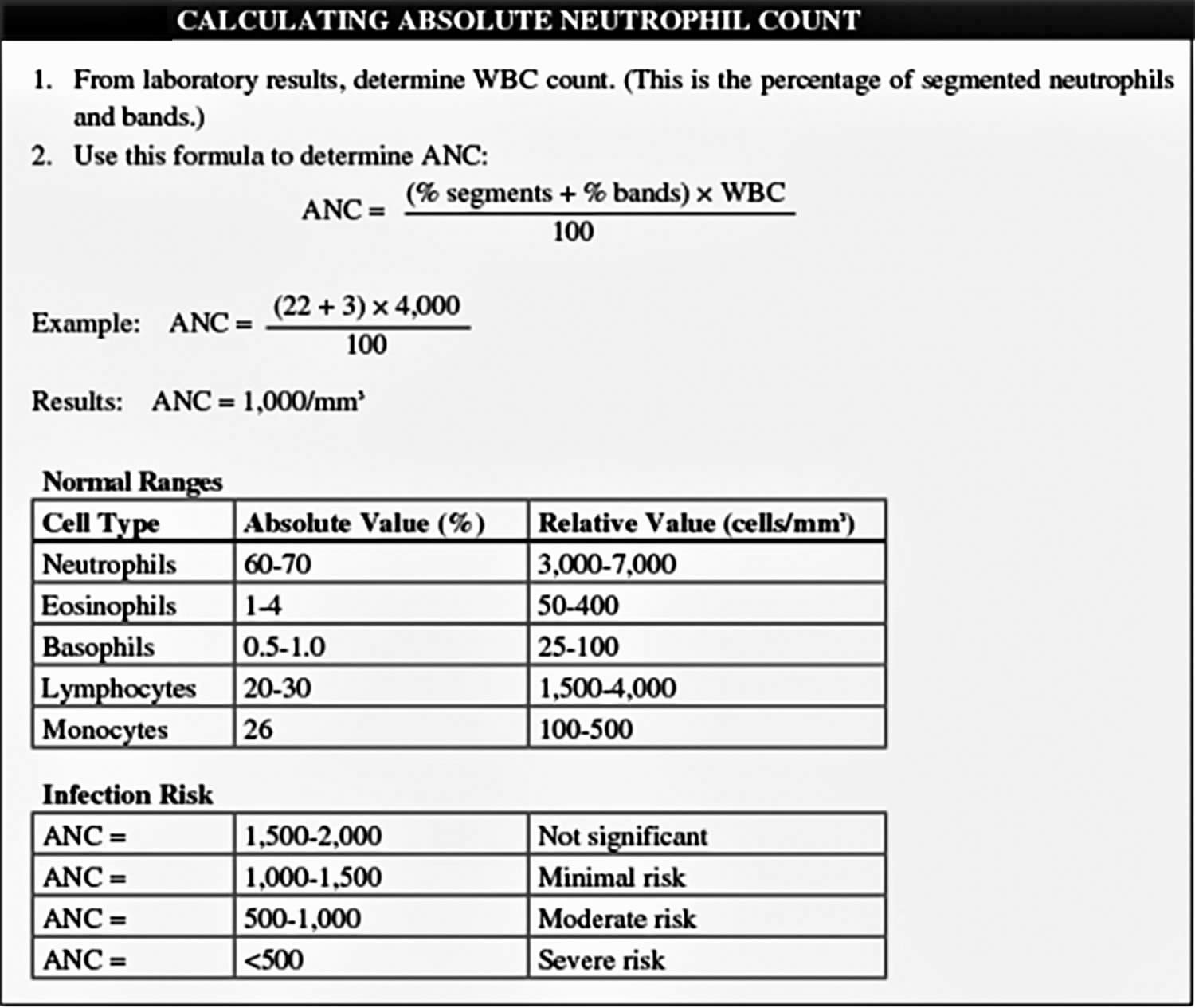
The absolute neutrophil count is found by multiplying the white blood cell count by the percent of neutrophils in the blood. For instance, if the white blood cell count is 8,000/mm³ and 50% of the white blood cells are neutrophils, the absolute neutrophil count is 4,000 (8,000 × 0.50 = 4,000).
When the absolute neutrophil count drops below 1,000/mm³ it is called neutropenia. Neutropenia refers to an abnormally low absolute neutrophil count. Your doctor will watch your absolute neutrophil count closely because the risk of infection is much higher when the absolute neutrophil count is below 500. A value above 1,000/mm³ usually means it’s safe to continue chemotherapy. On the other hand, a value below 1,000/mm³ sharply increases the risk of infection.
Uses of the absolute neutrophil count
Knowing a patient’s absolute neutrophil count has several advantages:
- It reflects the patient’s immunologic status in response to chemotherapy. Myelosuppressive chemotherapy suppresses white blood cell (and thus neutrophil) production, raising the risk of severe infection. A low absolute neutrophil count may indicate the need to delay the next chemotherapy dose or to reduce the dosage.
- The absolute neutrophil count value helps predict subsequent neutropenic events (such as fever) after the first chemotherapy cycle by providing a more accurate picture of immunologic status than the neutrophil or white blood cell count alone.
- It helps the physician determine whether to initiate treatment with growth factors, antibiotics, and other protective measures to decrease the risk of neutropenic complications. For example, administering filgrastim (recombinant methionyl human granulocyte colony-stimulating factor) can shorten neutropenia duration and reduce the risk of febrile neutropenia. This in turn increases the chance that full-dose chemotherapy can be given on time.
- For a hospitalized patient, a low absolute neutrophil count may indicate the need for protective isolation to guard against exposure to infection. For an outpatient, it may indicate the need to avoid crowds and people with colds, runny noses, or influenza.
How to calculate absolute neutrophil count
You can calculate your patient’s absolute neutrophil count in one of two ways.
Figure 9. How to calculate absolute neutrophil count (ANC)
Neutrophils low
Neutropenia is an abnormally low level of neutrophils. Neutrophils are a common type of white blood cell important to fighting off infections — particularly those caused by bacteria.
For adults, counts of less than 1,500 neutrophils per microliter of blood are considered to be neutropenia. For children, the cell count indicating neutropenia varies with age.
Some people have lower-than-average neutrophil counts, but not an increased risk of infection. In these situations their neutropenia isn’t a concern. Neutrophil counts less than 1,000 neutrophils per microliter — and especially counts of less than 500 neutrophils per microliter — are always considered to be neutropenia, where even the normal bacteria from your mouth and digestive tract can cause serious infections.
If you have neutropenia, you might have no symptoms at all. Some people get bacterial infections on different parts of the body like their skin, mouth area and gums, sinuses or internal organs such as their lungs.
Neutropenia is usually found when your doctor orders tests for a condition you’re already experiencing. It’s rare for neutropenia to be discovered unexpectedly or by chance.
Talk to your doctor about what your test results mean. Neutropenia and results from other tests might indicate the cause of your illness. Or, your doctor may suggest other tests to further check your condition.
Because neutropenia makes you vulnerable to bacterial and fungal infections, your doctor will probably advise certain precautions. These often include wearing a face mask, avoiding anyone with a cold, and washing your hands regularly and thoroughly.
Low neutrophils causes
Cancer chemotherapy is probably the most common cause of neutropenia. People with chemotherapy-related neutropenia are prone to infections while they wait for their cell counts to recover.
Neutrophils are manufactured in bone marrow — the spongy tissue inside some of your larger bones. Anything that disrupts neutrophil production can result in neutropenia.
Specific causes include:
- Chemotherapy
- Chronic idiopathic neutropenia in adults
- Kostmann’s syndrome (a congenital disorder involving low production of neutrophils)
- Leukemia and other diseases that damage bone marrow
- Myelodysplastic syndromes
- Myelofibrosis (a bone marrow disorder)
- Myelokathexis (a congenital disorder involving failure of neutrophils to enter the bloodstream)
- Radiation therapy
- Vitamin deficiencies
- Cancer that spreads to the bone marrow
- Aplastic anemia
Certain infections also can result in neutropenia, including:
- Hepatitis A
- Hepatitis B
- Hepatitis C
- HIV/AIDS
- Lyme disease
- Malaria
- Other viral infections that disrupt the work of bone marrow
- Salmonella infection
- Sepsis (an overwhelming bloodstream infection that uses up neutrophils faster than they can be produced)
Conditions that destroy neutrophils in the bloodstream and that can result in neutropenia include:
- Hypersplenism (an abnormality of the spleen causing blood cell destruction)
- Medications, such as antibiotics
- Rheumatoid arthritis and other autoimmune disorders
Neutropenia diagnosis and treatment
Neutropenia is diagnosed by a blood test. Other tests might be needed to find out the cause.
The treatment will depend on the cause of the neutropenia, and how severe it is.
If you have neutropenia, you should try to protect yourself against getting infections. You can do this by:
- washing your hands regularly
- avoiding large crowds
- wearing shoes all the time
- avoiding manicures and pedicures
- eating well and getting enough sleep
If you think you might have an infection, see you doctor quickly or go to the nearest emergency department.
Absolute neutrophils high
A high neutrophils count is an increase in disease-fighting cells in your blood. Also known as neutrophilia.
A high white blood cell count is usually found when your doctor orders tests to help diagnose a condition you’re already experiencing. It’s rarely an unexpected finding or simply discovered by chance.
Talk to your doctor about what these results mean. A high white blood cell count, along with results from other tests, might already indicate the cause of your illness. Or your doctor may suggest other tests to further evaluate your condition.
Common cause of neutrophilia are:
- Acute bacterial infections and also some infections caused by viruses and fungi
- Inflammation (e.g., inflammatory bowel disease, rheumatoid arthritis)
- Tissue death (necrosis) caused by trauma, major surgery, heart attack, burns
- Physiological (stress, rigorous exercise)
- Smoking
- Pregnancy—last trimester or during labor
- Chronic leukemia (e.g., myelogenous leukemia)
Specific causes of a high white blood cell count include:
- Acute lymphocytic leukemia
- Acute myelogenous leukemia (AML)
- Allergy, especially severe allergic reactions
- Chronic lymphocytic leukemia
- Chronic myelogenous leukemia
- Drugs, such as corticosteroids and epinephrine
- Infections, bacterial or viral
- Myelofibrosis (a bone marrow disorder)
- Polycythemia vera
- Rheumatoid arthritis (inflammatory joint disease)
- Stress, such as severe emotional or physical stress
- Tuberculosis
- Whooping cough
- Mócsai A. Diverse novel functions of neutrophils in immunity, inflammation, and beyond. The Journal of Experimental Medicine. 2013;210(7):1283-1299. doi:10.1084/jem.20122220. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3698517/
- Neutrophils and immunity: challenges and opportunities. Nathan C. Nat Rev Immunol. 2006 Mar; 6(3):173-82. https://www.ncbi.nlm.nih.gov/pubmed/16498448/
- The role of neutrophils in autoimmune diseases. Németh T, Mócsai A. Immunol Lett. 2012 Mar 30; 143(1):9-19. https://www.ncbi.nlm.nih.gov/pubmed/22342996/
- The production of cytokines by polymorphonuclear neutrophils. Cassatella MA. Immunol Today. 1995 Jan; 16(1):21-6.
- Breaching multiple barriers: leukocyte motility through venular walls and the interstitium. Nourshargh S, Hordijk PL, Sixt M. Nat Rev Mol Cell Biol. 2010 May; 11(5):366-78.
- Neutrophil granules: a library of innate immunity proteins. Borregaard N, Sørensen OE, Theilgaard-Mönch K. Trends Immunol. 2007 Aug; 28(8):340-5.
- Killing activity of neutrophils is mediated through activation of proteases by K+ flux. Reeves EP, Lu H, Jacobs HL, Messina CG, Bolsover S, Gabella G, Potma EO, Warley A, Roes J, Segal AW. Nature. 2002 Mar 21; 416(6878):291-7.
- Neutrophil extracellular traps kill bacteria. Brinkmann V, Reichard U, Goosmann C, Fauler B, Uhlemann Y, Weiss DS, Weinrauch Y, Zychlinsky A. Science. 2004 Mar 5; 303(5663):1532-5.
- Neutrophil function: from mechanisms to disease. Amulic B, Cazalet C, Hayes GL, Metzler KD, Zychlinsky A. Annu Rev Immunol. 2012; 30():459-89. https://www.ncbi.nlm.nih.gov/pubmed/22224774/
- Peculiarities of cell death mechanisms in neutrophils. Geering B, Simon HU. Cell Death Differ. 2011 Sep; 18(9):1457-69.
- In vivo labeling with 2H2O reveals a human neutrophil lifespan of 5.4 days. Pillay J, den Braber I, Vrisekoop N, Kwast LM, de Boer RJ, Borghans JA, Tesselaar K, Koenderman L. Blood. 2010 Jul 29; 116(4):625-7.
- Neutrophils in the activation and regulation of innate and adaptive immunity. Mantovani A, Cassatella MA, Costantini C, Jaillon S. Nat Rev Immunol. 2011 Jul 25; 11(8):519-31.
- Uribe-Querol E, Rosales C. Neutrophils in Cancer: Two Sides of the Same Coin. Journal of Immunology Research. 2015;2015:983698. doi:10.1155/2015/983698. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4706937/
- Phagosome dynamics during phagocytosis by neutrophils. Nordenfelt P, Tapper H. J Leukoc Biol. 2011 Aug; 90(2):271-84.
- Role of Nox family NADPH oxidases in host defense. Leto TL, Geiszt M. Antioxid Redox Signal. 2006 Sep-Oct; 8(9-10):1549-61.
- BK channels in innate immune functions of neutrophils and macrophages. Essin K, Gollasch M, Rolle S, Weissgerber P, Sausbier M, Bohn E, Autenrieth IB, Ruth P, Luft FC, Nauseef WM, Kettritz R. Blood. 2009 Feb 5; 113(6):1326-31.
- Infection-induced NETosis is a dynamic process involving neutrophil multitasking in vivo. Yipp BG, Petri B, Salina D, Jenne CN, Scott BN, Zbytnuik LD, Pittman K, Asaduzzaman M, Wu K, Meijndert HC, Malawista SE, de Boisfleury Chevance A, Zhang K, Conly J, Kubes P. Nat Med. 2012 Sep; 18(9):1386-93.
- Neutrophil extracellular traps: is immunity the second function of chromatin? Brinkmann V, Zychlinsky A. J Cell Biol. 2012 Sep 3; 198(5):773-83.
- Neutrophil clearance: when the party is over, clean-up begins. Bratton DL, Henson PM. Trends Immunol. 2011 Aug; 32(8):350-7.
- Phagocyte partnership during the onset and resolution of inflammation. Soehnlein O, Lindbom L. Nat Rev Immunol. 2010 Jun; 10(6):427-39.
- Role of neutrophil apoptosis in the resolution of inflammation. El Kebir D, Filep JG. ScientificWorldJournal. 2010 Sep 1; 10():1731-48.
- Neutrophil extracellular traps mediate a host defense response to human immunodeficiency virus-1. Saitoh T, Komano J, Saitoh Y, Misawa T, Takahama M, Kozaki T, Uehata T, Iwasaki H, Omori H, Yamaoka S, Yamamoto N, Akira S. Cell Host Microbe. 2012 Jul 19; 12(1):109-16.
- Neutrophils recruited to sites of infection protect from virus challenge by releasing neutrophil extracellular traps. Jenne CN, Wong CH, Zemp FJ, McDonald B, Rahman MM, Forsyth PA, McFadden G, Kubes P. Cell Host Microbe. 2013 Feb 13; 13(2):169-80.
- Endocytosis of soluble immune complexes leads to their clearance by FcγRIIIB but induces neutrophil extracellular traps via FcγRIIA in vivo. Chen K, Nishi H, Travers R, Tsuboi N, Martinod K, Wagner DD, Stan R, Croce K, Mayadas TN. Blood. 2012 Nov 22; 120(22):4421-31.
- Neutrophils transport antigen from the dermis to the bone marrow, initiating a source of memory CD8+ T cells. Duffy D, Perrin H, Abadie V, Benhabiles N, Boissonnas A, Liard C, Descours B, Reboulleau D, Bonduelle O, Verrier B, Van Rooijen N, Combadière C, Combadière B. Immunity. 2012 Nov 16; 37(5):917-29.
- Neutrophils in tuberculosis: friend or foe? Lowe DM, Redford PS, Wilkinson RJ, O’Garra A, Martineau AR. Trends Immunol. 2012 Jan; 33(1):14-25.
- Neutrophil-mediated innate immune resistance to mycobacteria. Martineau AR, Newton SM, Wilkinson KA, Kampmann B, Hall BM, Nawroly N, Packe GE, Davidson RN, Griffiths CJ, Wilkinson RJ. J Clin Invest. 2007 Jul; 117(7):1988-94.
- An interferon-inducible neutrophil-driven blood transcriptional signature in human tuberculosis. Berry MP, Graham CM, McNab FW, Xu Z, Bloch SA, Oni T, Wilkinson KA, Banchereau R, Skinner J, Wilkinson RJ, Quinn C, Blankenship D, Dhawan R, Cush JJ, Mejias A, Ramilo O, Kon OM, Pascual V, Banchereau J, Chaussabel D, O’Garra A. Nature. 2010 Aug 19; 466(7309):973-7.
- Neutrophils rapidly migrate via lymphatics after Mycobacterium bovis BCG intradermal vaccination and shuttle live bacilli to the draining lymph nodes. Abadie V, Badell E, Douillard P, Ensergueix D, Leenen PJ, Tanguy M, Fiette L, Saeland S, Gicquel B, Winter N. Blood. 2005 Sep 1; 106(5):1843-50.
- Immunology. Neutrophil soldiers or Trojan Horses? John B, Hunter CA. Science. 2008 Aug 15; 321(5891):917-8.
- Pathological role of interleukin 17 in mice subjected to repeated BCG vaccination after infection with Mycobacterium tuberculosis. Cruz A, Fraga AG, Fountain JJ, Rangel-Moreno J, Torrado E, Saraiva M, Pereira DR, Randall TD, Pedrosa J, Cooper AM, Castro AG. J Exp Med. 2010 Aug 2; 207(8):1609-16.
- Regulation of neutrophils by interferon-γ limits lung inflammation during tuberculosis infection. Nandi B, Behar SM. J Exp Med. 2011 Oct 24; 208(11):2251-62.
- Coactivation of Syk kinase and MyD88 adaptor protein pathways by bacteria promotes regulatory properties of neutrophils. Zhang X, Majlessi L, Deriaud E, Leclerc C, Lo-Man R. Immunity. 2009 Nov 20; 31(5):761-71.
- Failure to detect production of IL-10 by activated human neutrophils. Davey MS, Tamassia N, Rossato M, Bazzoni F, Calzetti F, Bruderek K, Sironi M, Zimmer L, Bottazzi B, Mantovani A, Brandau S, Moser B, Eberl M, Cassatella MA. Nat Immunol. 2011 Oct 19; 12(11):1017-8; author reply 1018-20.
- Neutrophils exert protection in the early tuberculous granuloma by oxidative killing of mycobacteria phagocytosed from infected macrophages. Yang CT, Cambier CJ, Davis JM, Hall CJ, Crosier PS, Ramakrishnan L. Cell Host Microbe. 2012 Sep 13; 12(3):301-12.
- Critical role of the neutrophil-associated high-affinity receptor for IgE in the pathogenesis of experimental cerebral malaria. Porcherie A, Mathieu C, Peronet R, Schneider E, Claver J, Commere PH, Kiefer-Biasizzo H, Karasuyama H, Milon G, Dy M, Kinet JP, Louis J, Blank U, Mécheri S. J Exp Med. 2011 Oct 24; 208(11):2225-36.
- Blood dendritic cells interact with splenic marginal zone B cells to initiate T-independent immune responses. Balázs M, Martin F, Zhou T, Kearney J. Immunity. 2002 Sep; 17(3):341-52.
- G-CSF-stimulated neutrophils are a prominent source of functional BLyS. Scapini P, Nardelli B, Nadali G, Calzetti F, Pizzolo G, Montecucco C, Cassatella MA. J Exp Med. 2003 Feb 3; 197(3):297-302.
- Neutrophil-derived APRIL concentrated in tumor lesions by proteoglycans correlates with human B-cell lymphoma aggressiveness. Schwaller J, Schneider P, Mhawech-Fauceglia P, McKee T, Myit S, Matthes T, Tschopp J, Donze O, Le Gal FA, Huard B. Blood. 2007 Jan 1; 109(1):331-8.
- APRIL secreted by neutrophils binds to heparan sulfate proteoglycans to create plasma cell niches in human mucosa. Huard B, McKee T, Bosshard C, Durual S, Matthes T, Myit S, Donze O, Frossard C, Chizzolini C, Favre C, Zubler R, Guyot JP, Schneider P, Roosnek E. J Clin Invest. 2008 Aug; 118(8):2887-95.
- B cell-helper neutrophils stimulate the diversification and production of immunoglobulin in the marginal zone of the spleen. Puga I, Cols M, Barra CM, He B, Cassis L, Gentile M, Comerma L, Chorny A, Shan M, Xu W, Magri G, Knowles DM, Tam W, Chiu A, Bussel JB, Serrano S, Lorente JA, Bellosillo B, Lloreta J, Juanpere N, Alameda F, Baró T, de Heredia CD, Torán N, Català A, Torrebadell M, Fortuny C, Cusí V, Carreras C, Diaz GA, Blander JM, Farber CM, Silvestri G, Cunningham-Rundles C, Calvillo M, Dufour C, Notarangelo LD, Lougaris V, Plebani A, Casanova JL, Ganal SC, Diefenbach A, Aróstegui JI, Juan M, Yagüe J, Mahlaoui N, Donadieu J, Chen K, Cerutti A. Nat Immunol. 2011 Dec 25; 13(2):170-80.
- CCR7 is involved in the migration of neutrophils to lymph nodes. Beauvillain C, Cunin P, Doni A, Scotet M, Jaillon S, Loiry ML, Magistrelli G, Masternak K, Chevailler A, Delneste Y, Jeannin P. Blood. 2011 Jan 27; 117(4):1196-204.
- Neutrophils efficiently cross-prime naive T cells in vivo. Beauvillain C, Delneste Y, Scotet M, Peres A, Gascan H, Guermonprez P, Barnaba V, Jeannin P. Blood. 2007 Oct 15; 110(8):2965-73.
- Mouse neutrophils are professional antigen-presenting cells programmed to instruct Th1 and Th17 T-cell differentiation. Abi Abdallah DS, Egan CE, Butcher BA, Denkers EY. Int Immunol. 2011 May; 23(5):317-26.
- A subset of neutrophils in human systemic inflammation inhibits T cell responses through Mac-1. Pillay J, Kamp VM, van Hoffen E, Visser T, Tak T, Lammers JW, Ulfman LH, Leenen LP, Pickkers P, Koenderman L. J Clin Invest. 2012 Jan; 122(1):327-36.
- Neutrophils control the magnitude and spread of the immune response in a thromboxane A2-mediated process. Yang CW, Unanue ER. J Exp Med. 2013 Feb 11; 210(2):375-87.
- Myeloid-derived suppressor cells as regulators of the immune system. Gabrilovich DI, Nagaraj S. Nat Rev Immunol. 2009 Mar; 9(3):162-74.
- Neutrophil-derived CCL3 is essential for the rapid recruitment of dendritic cells to the site of Leishmania major inoculation in resistant mice. Charmoy M, Brunner-Agten S, Aebischer D, Auderset F, Launois P, Milon G, Proudfoot AE, Tacchini-Cottier F. PLoS Pathog. 2010 Feb 5; 6(2):e1000755.
- Polymorphonuclear neutrophils deliver activation signals and antigenic molecules to dendritic cells: a new link between leukocytes upstream of T lymphocytes. Megiovanni AM, Sanchez F, Robledo-Sarmiento M, Morel C, Gluckman JC, Boudaly S. J Leukoc Biol. 2006 May; 79(5):977-88.
- Alarmins link neutrophils and dendritic cells. Yang D, de la Rosa G, Tewary P, Oppenheim JJ. Trends Immunol. 2009 Nov; 30(11):531-7.
- The interferon-alpha signature of systemic lupus erythematosus. Obermoser G, Pascual V. Lupus. 2010 Aug; 19(9):1012-9.
- Neutrophils in the pathogenesis and manifestations of SLE. Kaplan MJ. Nat Rev Rheumatol. 2011 Sep 27; 7(12):691-9.
- Neutrophils activate plasmacytoid dendritic cells by releasing self-DNA-peptide complexes in systemic lupus erythematosus. Lande R, Ganguly D, Facchinetti V, Frasca L, Conrad C, Gregorio J, Meller S, Chamilos G, Sebasigari R, Riccieri V, Bassett R, Amuro H, Fukuhara S, Ito T, Liu YJ, Gilliet M. Sci Transl Med. 2011 Mar 9; 3(73):73ra19.
- Fc gamma RIIa is expressed on natural IFN-alpha-producing cells (plasmacytoid dendritic cells) and is required for the IFN-alpha production induced by apoptotic cells combined with lupus IgG. Båve U, Magnusson M, Eloranta ML, Perers A, Alm GV, Rönnblom L. J Immunol. 2003 Sep 15; 171(6):3296-302.
- Netting neutrophils are major inducers of type I IFN production in pediatric systemic lupus erythematosus. Garcia-Romo GS, Caielli S, Vega B, Connolly J, Allantaz F, Xu Z, Punaro M, Baisch J, Guiducci C, Coffman RL, Barrat FJ, Banchereau J, Pascual V. Sci Transl Med. 2011 Mar 9; 3(73):73ra20.
- Mutation of DNASE1 in people with systemic lupus erythematosus. Yasutomo K, Horiuchi T, Kagami S, Tsukamoto H, Hashimura C, Urushihara M, Kuroda Y. Nat Genet. 2001 Aug; 28(4):313-4.
- Netting neutrophils in autoimmune small-vessel vasculitis. Kessenbrock K, Krumbholz M, Schönermarck U, Back W, Gross WL, Werb Z, Gröne HJ, Brinkmann V, Jenne DE. Nat Med. 2009 Jun; 15(6):623-5.
- Crosstalk between neutrophils, B-1a cells and plasmacytoid dendritic cells initiates autoimmune diabetes. Diana J, Simoni Y, Furio L, Beaudoin L, Agerberth B, Barrat F, Lehuen A. Nat Med. 2013 Jan; 19(1):65-73.
- Novel cell death program leads to neutrophil extracellular traps. Fuchs TA, Abed U, Goosmann C, Hurwitz R, Schulze I, Wahn V, Weinrauch Y, Brinkmann V, Zychlinsky A. J Cell Biol. 2007 Jan 15; 176(2):231-41.
- NADPH oxidase inhibits the pathogenesis of systemic lupus erythematosus. Campbell AM, Kashgarian M, Shlomchik MJ. Sci Transl Med. 2012 Oct 24; 4(157):157ra141.
- Fc receptors and their role in immune regulation and autoimmunity. Takai T. J Clin Immunol. 2005 Jan; 25(1):1-18.
- The defensive alliance between neutrophils and NK cells as a novel arm of innate immunity. Costantini C, Cassatella MA. J Leukoc Biol. 2011 Feb; 89(2):221-33.
- A novel role for neutrophils as critical activators of NK cells. Spörri R, Joller N, Hilbi H, Oxenius A. J Immunol. 2008 Nov 15; 181(10):7121-30.
- Neutrophil depletion impairs natural killer cell maturation, function, and homeostasis. Jaeger BN, Donadieu J, Cognet C, Bernat C, Ordoñez-Rueda D, Barlogis V, Mahlaoui N, Fenis A, Narni-Mancinelli E, Beaupain B, Bellanné-Chantelot C, Bajénoff M, Malissen B, Malissen M, Vivier E, Ugolini S. J Exp Med. 2012 Mar 12; 209(3):565-80.
- Inflammatory reactions and severe neutropenia in mice lacking the transcriptional repressor Gfi1. Karsunky H, Zeng H, Schmidt T, Zevnik B, Kluge R, Schmid KW, Dührsen U, Möröy T. Nat Genet. 2002 Mar; 30(3):295-300.
- Mutations in proto-oncogene GFI1 cause human neutropenia and target ELA2. Person RE, Li FQ, Duan Z, Benson KF, Wechsler J, Papadaki HA, Eliopoulos G, Kaufman C, Bertolone SJ, Nakamoto B, Papayannopoulou T, Grimes HL, Horwitz M. Nat Genet. 2003 Jul; 34(3):308-12.
- Human neutrophils interact with both 6-sulfo LacNAc+ DC and NK cells to am plify NK-derived IFN{gamma}: role of CD18, ICAM-1, and ICAM-3. Costantini C, Calzetti F, Perbellini O, Micheletti A, Scarponi C, Lonardi S, Pelletier M, Schakel K, Pizzolo G, Facchetti F, Vermi W, Albanesi C, Cassatella MA. Blood. 2011 Feb 3; 117(5):1677-86.
- Pathophysiology of anaphylaxis. Khan BQ, Kemp SF. Curr Opin Allergy Clin Immunol. 2011 Aug; 11(4):319-25.
- Basophils orchestrate chronic allergic dermatitis and protective immunity against helminths. Ohnmacht C, Schwartz C, Panzer M, Schiedewitz I, Naumann R, Voehringer D. Immunity. 2010 Sep 24; 33(3):364-74.
- Mouse and human neutrophils induce anaphylaxis. Jönsson F, Mancardi DA, Kita Y, Karasuyama H, Iannascoli B, Van Rooijen N, Shimizu T, Daëron M, Bruhns P. J Clin Invest. 2011 Apr; 121(4):1484-96.
- Inflammation, atherosclerosis, and coronary artery disease. Hansson GK. N Engl J Med. 2005 Apr 21; 352(16):1685-95.
- Distinct infiltration of neutrophils in lesion shoulders in ApoE-/- mice. Rotzius P, Thams S, Soehnlein O, Kenne E, Tseng CN, Björkström NK, Malmberg KJ, Lindbom L, Eriksson EE. Am J Pathol. 2010 Jul; 177(1):493-500.
- High neutrophil numbers in human carotid atherosclerotic plaques are associated with characteristics of rupture-prone lesions. Ionita MG, van den Borne P, Catanzariti LM, Moll FL, de Vries JP, Pasterkamp G, Vink A, de Kleijn DP. Arterioscler Thromb Vasc Biol. 2010 Sep; 30(9):1842-8.
- Hyperlipidemia-triggered neutrophilia promotes early atherosclerosis. Drechsler M, Megens RT, van Zandvoort M, Weber C, Soehnlein O. Circulation. 2010 Nov 2; 122(18):1837-45.
- Lack of neutrophil-derived CRAMP reduces atherosclerosis in mice. Döring Y, Drechsler M, Wantha S, Kemmerich K, Lievens D, Vijayan S, Gallo RL, Weber C, Soehnlein O. Circ Res. 2012 Apr 13; 110(8):1052-6.
- Auto-antigenic protein-DNA complexes stimulate plasmacytoid dendritic cells to promote atherosclerosis. Döring Y, Manthey HD, Drechsler M, Lievens D, Megens RT, Soehnlein O, Busch M, Manca M, Koenen RR, Pelisek J, Daemen MJ, Lutgens E, Zenke M, Binder CJ, Weber C, Zernecke A. Circulation. 2012 Apr 3; 125(13):1673-83.
- Neutrophils mediate insulin resistance in mice fed a high-fat diet through secreted elastase. Talukdar S, Oh DY, Bandyopadhyay G, Li D, Xu J, McNelis J, Lu M, Li P, Yan Q, Zhu Y, Ofrecio J, Lin M, Brenner MB, Olefsky JM. Nat Med. 2012 Sep; 18(9):1407-12.
- Ccr5 but not Ccr1 deficiency reduces development of diet-induced atherosclerosis in mice. Braunersreuther V, Zernecke A, Arnaud C, Liehn EA, Steffens S, Shagdarsuren E, Bidzhekov K, Burger F, Pelli G, Luckow B, Mach F, Weber C. Arterioscler Thromb Vasc Biol. 2007 Feb; 27(2):373-9.
- Circulating activated platelets exacerbate atherosclerosis in mice deficient in apolipoprotein E. Huo Y, Schober A, Forlow SB, Smith DF, Hyman MC, Jung S, Littman DR, Weber C, Ley K. Nat Med. 2003 Jan; 9(1):61-7.
- P-Selectin or intercellular adhesion molecule (ICAM)-1 deficiency substantially protects against atherosclerosis in apolipoprotein E-deficient mice. Collins RG, Velji R, Guevara NV, Hicks MJ, Chan L, Beaudet AL. J Exp Med. 2000 Jan 3; 191(1):189-94.





{kind=link}