Quadrantanopia
Quadrantanopia also called quadrantanopsia, refers to a defect in the visual field affecting a quarter of the field of vision. Quadrantanopia can be associated with a lesion of optic tract pathway. While quadrantanopia can be caused by lesions in the temporal and parietal lobes, it is most commonly associated with lesions in the occipital lobe 1.
An interesting aspect of quadrantanopia is that there exists a distinct and sharp border between the intact and damaged visual fields, due to an anatomical separation of the quadrants of the visual field. For example, information in the left half of visual field is processed in the right occipital lobe and information in the right half of the visual field is processed in the left occipital lobe 1.
In a quadrantanopia that is partial, there also exists a distinct and sharp border between the intact and damaged field within the quadrant. The sufferer is able to detect light within the damaged visual field.
The prospects of recovering vision in the affected field are bleak. Occasionally, patients will spontaneously recover vision in the affected field within the first three months after the brain injury; however, vision loss remaining after this period of spontaneous recovery is traditionally thought to be permanent, certain companies now claim to be able to induce recovery of vision after this three-month period.
Figure 1. Quadrantanopia

Homonymous quadrantanopia
Homonymous implies the defect does not cross the vertical meridian. Homonymous describes defects that affect the same side of the vertical meridian (i.e., right or left side) of both eyes.
Homonymous inferior quadrantanopia is a loss of vision in the same lower quadrant of visual field in both eyes whereas a homonymous superior quadrantanopia is a loss of vision in the same upper quadrant of visual field in both eyes 2.
A lesion affecting one side of the temporal lobe may cause damage to the inferior optic radiations (known as the temporal pathway or Meyer’s loop) which can lead to superior quadrantanopia on the contralateral side of both eyes (colloquially referred to as “pie in the sky”); if the superior optic radiations (parietal pathway) are lesioned, the visual loss occurs on the inferior contralateral side of both eyes and is referred to as an inferior quadrantanopia.
Binasal or bitemporal quadrantanopia
Binasal (either inferior or superior) quadrantanopia affects either the upper or lower inner visual quadrants closer to the nasal cavity in both eyes. Bitemporal (either inferior or superior) quadrantanopia affects either the upper or lower outer visual quadrants in both eyes.
Visual pathway anatomy
The optic nerve is the output of the retina. Therefore, each optic nerve carries all of the visual information from the eye from which it emerges. Since the left hemisphere moves the right side of the body and the right hemisphere moves the left side of the body, it makes sense that the left hemisphere should receive the visual information from the right half of the world and the right hemisphere should receive the visual information from the left half of the world. Therefore, some of the information in each optic nerve must cross so that the brain can work with the left and right visual fields rather than merely what is seen by the left and right eyes: The left hemisphere must receive right visual field information from both the left eye and the right eye; the right hemisphere must receive left visual field information from both the left eye and the right eye. The crossing of visual field information to convert the visual world from left eye–right eye organization to left field–right field organization occurs at the optic chiasm. Posterior to the optic chiasm, visual information is organized into fields: the left visual field is processed in the right hemisphere, and the right visual field in the left hemisphere.
The right visual field (blue) projects onto the medial (nasal) retina in the right eye and the lateral (temporal) retina in the left eye (Figures 3 to 5). The left visual field (tan) projects to the medial (nasal) retina in the left eye and the lateral (temporal) retina in the right eye. The medial (nasal) retinas are, therefore, receiving the lateral (temporal) visual fields, and the lateral (temporal) retinas are receiving the medial (nasal) visual fields.
All visual information from the right visual field must end up in the left hemisphere, and all visual information from the left visual field must end up in the right hemisphere. The right visual field information in the left optic nerve (from the lateral retina) is already on the correct (left) side of the brain, but the right visual field information in the right optic nerve (from the medial retina) must cross from the right optic nerve to the left side of the brain to join it. The left visual field information in the right optic nerve (from the lateral retina) is already on the correct side.
Optic nerve, optic chiasma and optic tract
- Optic nerve is made up of axons of ganglion cells of retina
- Fibers of the nerve arising from the four quadrants of retina maintain the same relative position within the nerve
- Fibers from the macula are numerous and form papillomacular bundle
- The fibers of the nasal half of each retina enter the optic tract of the opposite side after crossing in the optic chiasma
- Fibers from the temporal half enter the optic tract of same side
- Optic tract carries these fibers to lateral geniculate body of the corresponding side.
The axons in the optic tract terminate in four nuclei within the brain:
- the lateral geniculate nucleus of the thalamus – for visual perception;
- the superior colliculus of the midbrain – for control of eye movements;
- the pretectum of the midbrain – for control of the pupillary light reflex; and
- the suprachiasmatic nucleus of the hypothalamus – for control of diurnal rhythms and hormonal changes.
Lateral geniculate body (LGN)
- Part of metathalamus
- Grey matter is split into 6 layers
- Fibers from the same side eye end in laminae 2, 3 and 5.
- Fibers from opposite side eye end in 1, 4 and 6
- Macular fiber end in central and posterior part of body & this area is relatively large
The vast majority of optic tract fibers terminate on neurons in the lateral geniculate nucleus of the thalamus (Figure 5). Like the retina, the lateral geniculate nucleus is a laminated structure, in this case, with six principal layers of cells (Figure 5B).
- The largest cells form the deepest two (magnocellular) layers
- Smaller cells form the upper four (parvocellular) layers
- Thin layers of the smallest cells (i.e., the koniocellular neurons) are interposed between these principal layers.
The optic tract fibers from each eye synapse in different layers of the lateral geniculate nucleus. Consequently, each lateral geniculate nucleus neuron responds to stimulation of one eye only.
The functional properties of lateral geniculate nucleus neurons are similar to those of retinal ganglion cells.
The lateral geniculate nucleus neurons are monocular (i.e., respond to stimulation of one eye only) and have concentric (center-surround) receptive fields. The lateral geniculate nucleus neurons are segregated into three major groups:
- The neurons in the magnocellular layers (mlateral geniculate nucleus cells)
- process M-retinal ganglion cell inputs
- behave like M-retinal ganglion cells
- have relatively large center-surround receptive fields
- are color insensitive
- are most sensitive to movement of visual stimuli
- The neurons in the parvocellular layers (plateral geniculate nucleus cells)
- process P-retinal ganglion cell inputs
- behave like P-retinal ganglion cells
- have relatively small center-surround receptive fields
- are color sensitive
- are well suited for detecting contrasts that form the basis for shape/form discrimination.
- A third group, the koniocellular neurons (klateral geniculate nucleus)
- process P-retinal ganglion cell inputs
- behave like P-retinal ganglion cells
- have the smallest concentric receptive fields
- have stronger color sensitivity than P-retinal ganglion cells
- are well suited for detecting colors that aid in shape/form discrimination.
The axons of these different types of lateral geniculate nucleus neurons terminate in different layers or sublayers of the primary visual cortex.
Visual cortical areas
The primary visual cortical receiving area is in the occipital lobe. The primary visual cortex is characterized by a unique layered appearance in Nissl stained tissue. Consequently, it is called the striate cortex. It includes the calcarine cortex, which straddles the calcarine fissure, and extends around the occipital pole to include the lateral aspect of the caudal occipital lobe (Area 17).
The lateral geniculate nucleus neurons send their axons in the internal capsule to the occipital lobe where they terminate in the striate cortex (Figure 2).
- The lateral geniculate nucleus axons fan out as the optic radiations of the internal capsule and travel through the temporal, parietal and occipital lobes.
- The lateral geniculate nucleus axons in the sublenticular segment of the optic radiations pass below the lenticular nuclei, loop around the inferior horn of the lateral ventricle within the temporal lobe and swing posteriorly to form Meyer’s loop.
- Once around the inferior horn, they travel up to the inferior bank of the striate cortex, where they terminate.
- The lateral geniculate nucleus axons in the retrolenticular segment of the internal capsule pass superiorly through the parietal lobe to end in the superior bank of the striate cortex.
The striate cortex is considered to be the primary visual cortex or V1, as:
- most lateral geniculate nucleus axons terminate in V1
- all V1 neurons respond to visual stimuli exclusively
- ablating V1 results in blindness
- electrical stimulation of V1 elicits visual sensations.
The striate cortex is involved in the initial cortical processing of all visual information necessary for visual perception and its damage results in loss of vision in the contralesional hemifield.
The color, shape and movement information from the thalamus are sent to different neurons within V1 for further processing in V1 and then sent onto different areas of the extrastriate visual cortex.
Figure 2. Visual pathway
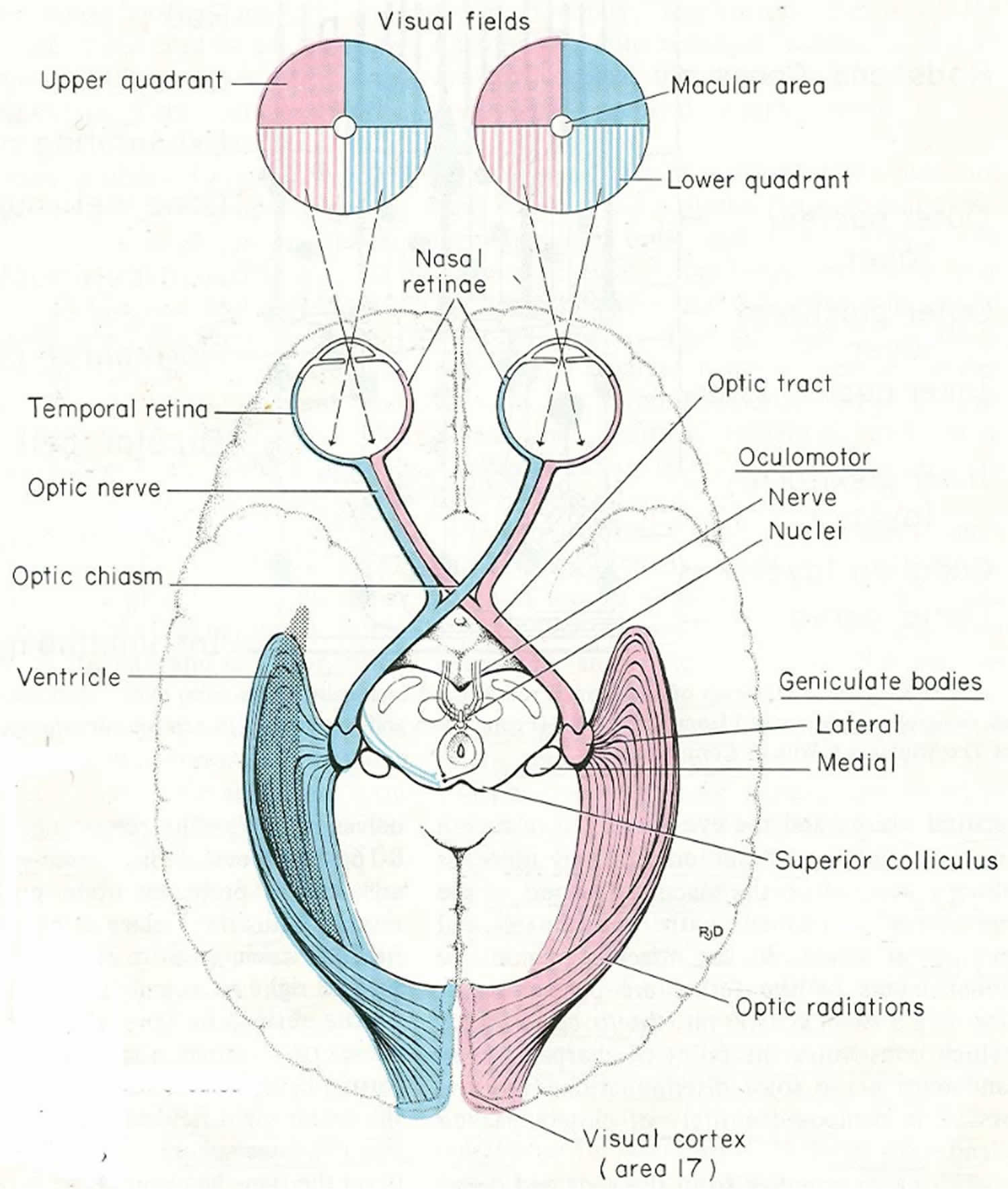
Figure 3. Schematic of visual pathway and visual field deficits caused by lesions in the visual pathway
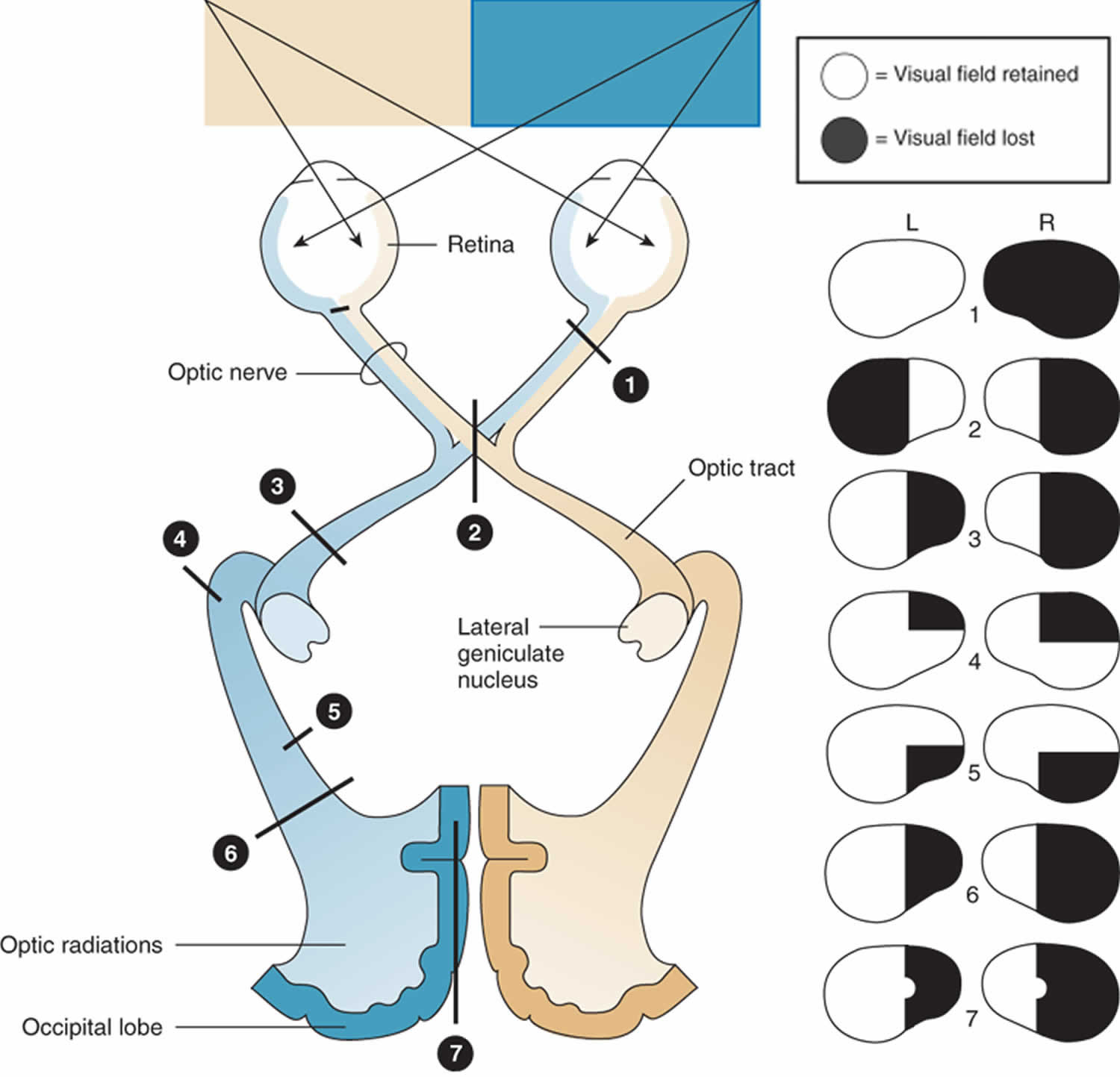
Figure 4. Visual field defects
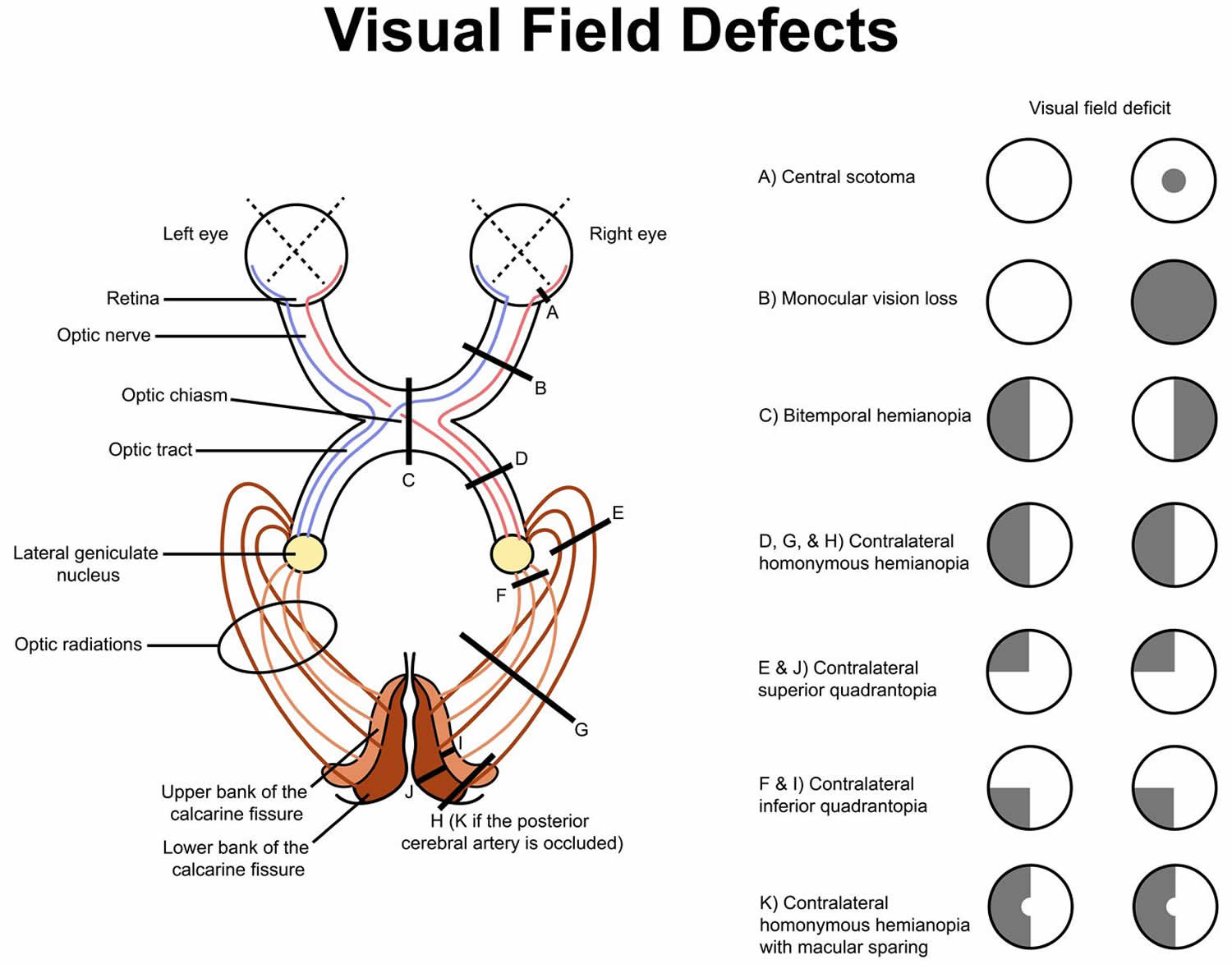 Figure 5. Lateral geniculate body
Figure 5. Lateral geniculate body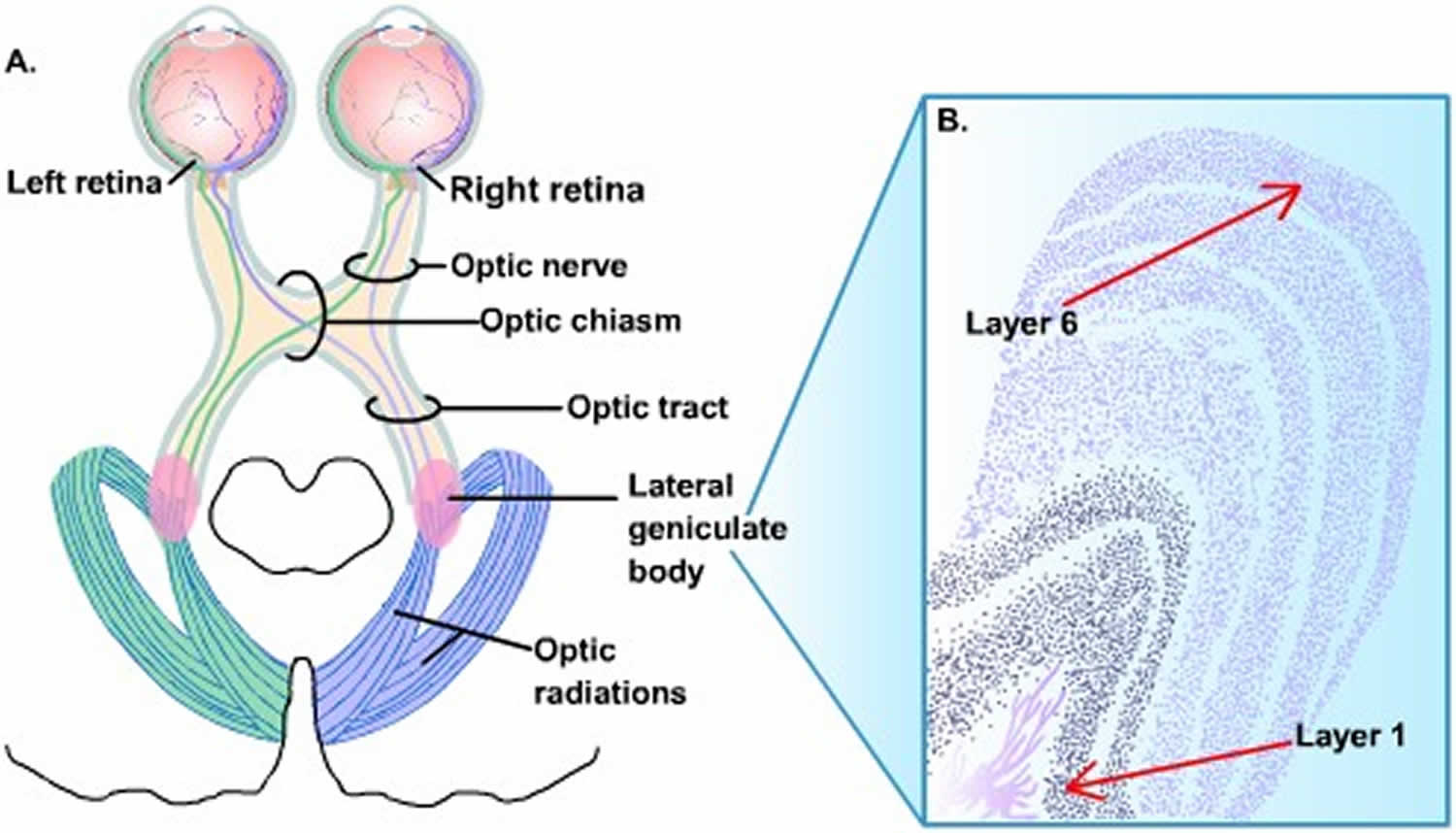
Quadrantanopia causes
Contralateral superior quadrantanopia (“pie in the sky”) causes:
- Temporal lobe lesions
- Lesions involving the lower bank of the calcarine fissure
Contralateral inferior quadrantanopia (“pie on the floor”) causes
- Parietal lobe lesions such as infarction of the superior division of the middle cerebral artery
- Lesions involving the superior bank of the calcarine fissure





{kind=link}