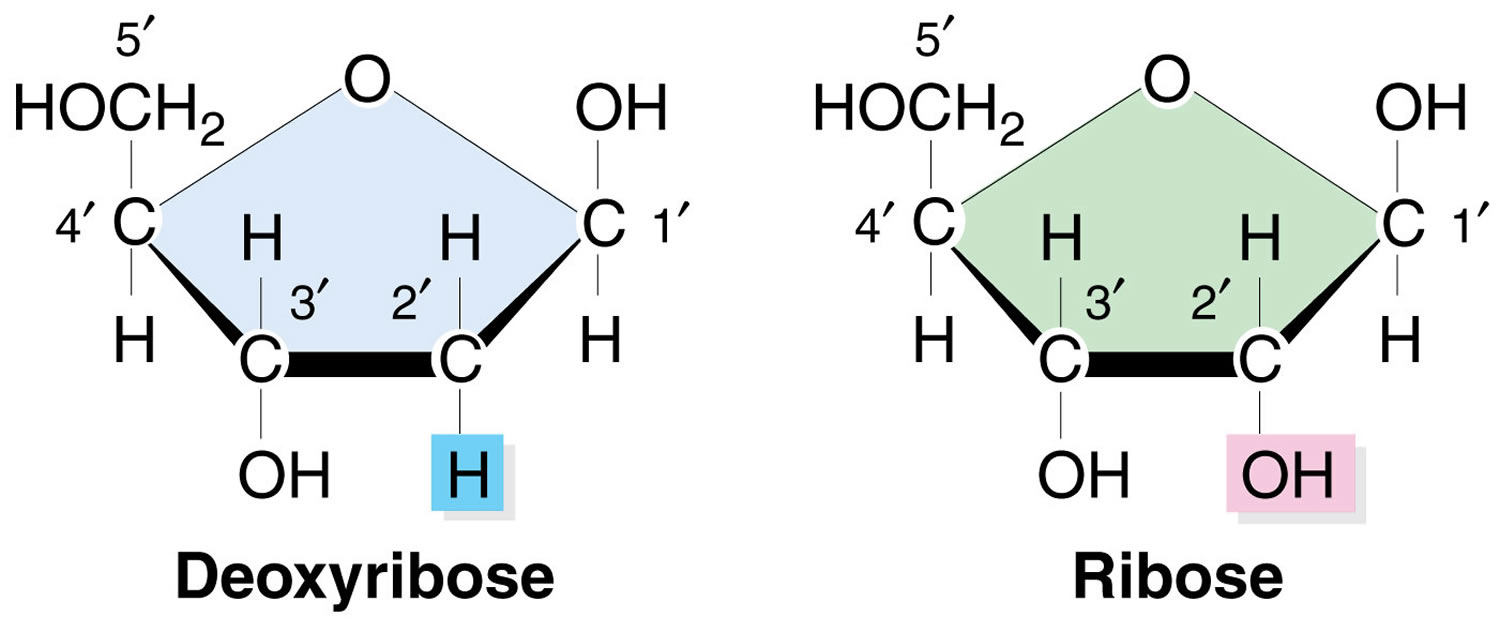
What is ribose
Ribose or D-Ribose is a naturally occurring pentose monosaccharide present in all living cells including the blood and is an essential component for biological energy production 1. D-Ribose is also a product of dietary nucleic acid digestion 2. Ribose or D-Ribose is a key component of many important biomolecules including ribonucleic acid (RNA) 3, adenosine triphosphate (ATP) 4 and Riboflavin (vitamin B2) 5. Many foods, such as wheat bran, eggs, meat, cheese and yeast, contain reasonable high concentrations of ribonucleic acid (RNA) and Riboflavin (vitamin B2). The pentose phosphate pathway can convert hexose to D-Ribose. Besides food, D-Ribose is orally administered to improve athletic performance and the ability to exercise by boosting muscle energy as a readily available source of energy.
Previous studies indicated that ribose is a promising supplement in patients with advanced heart failure in improving heart and skeletal muscle performances 6. Ribose has shown to replenish low myocardial energy levels, improving cardiac dysfunction following ischemia, and improving ventilation efficiency in patients with heart failure 6.
Figure 1. Ribose and D-ribose
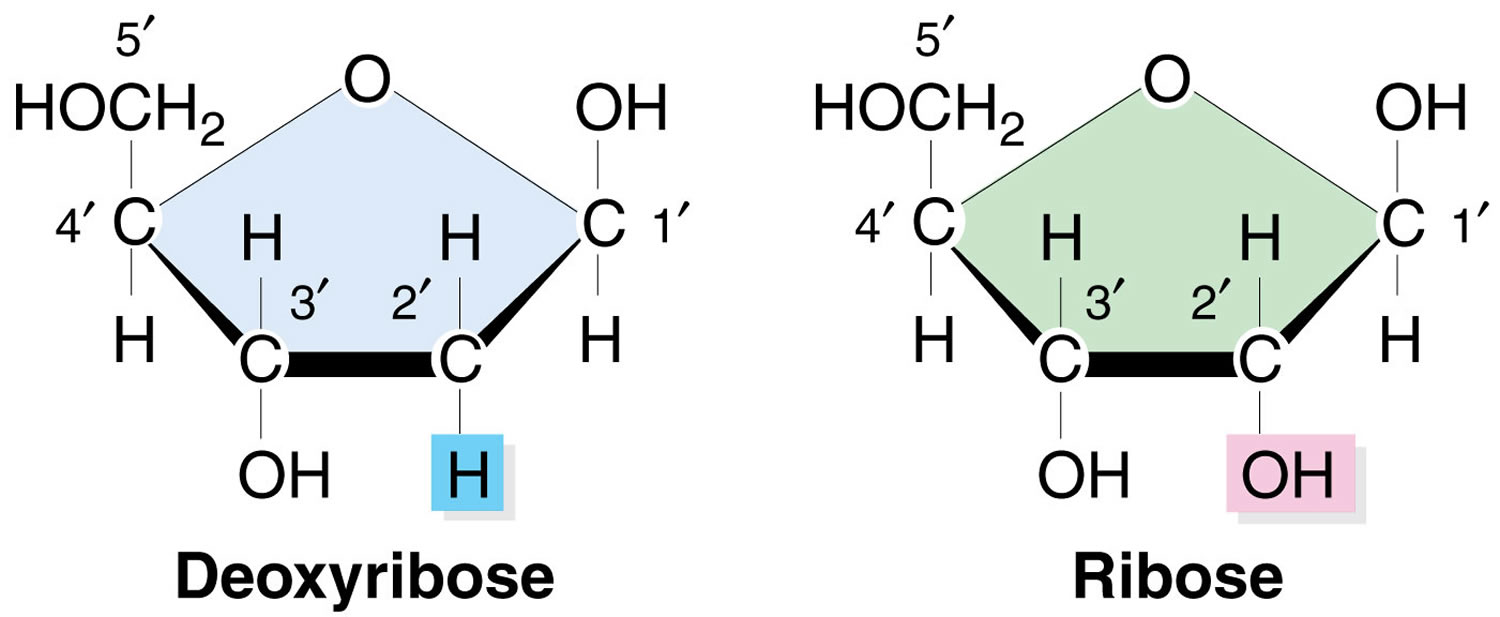
Footnote: Note the absence of the hydroxyl (-OH) group on the 2’ carbon in the deoxy-ribose sugar in DNA as compared with the ribose sugar in RNA.
D-Ribose is a popular dietary supplement for humans and horses because of its crucial role in cellular bioenergetics. It is also used to improve symptoms of diseases such as chronic fatigue syndrome, fibromyalgia 7 and coronary artery disease 8.
The available data indicate that in humans, D-Ribose is rapidly and nearly completely absorbed when administered at 200 mg/kg body weight per hour for 5 hours 9. In humans, at dose levels above 3 g (about 40 mg/kg body weight), absorption was faster than metabolism. Application of D-Ribose with meals decreases absorption. In the body, D-Ribose is converted mainly to glucose via the pentose phosphate pathway, rather than nucleic acid precursors, which then is further used in the metabolism/biosynthesis. Part of the ribose and its metabolites is excreted via the urine and the percentage increases with increasing dose.
Several human studies which were not designed to assess the safety of D-Ribose found the administration of single oral doses of 2–87 g D-Ribose consistently report transient decreases of glucose concentrations within 1–3 hours. The transient decrease of glucose concentration was not associated with clinical symptoms of hypoglycaemia, except for one case, where a low‐weight female experienced short‐term symptoms of hypoglycaemia after ingesting 10 g of ribose in the fasted state 9. While in this and in most of the other studies, blood glucose levels did not fall below 2.8 mmol/L, a temporary significant decline of blood glucose to 2.6 mmol/L was observed in one study following a single oral dose of 10 g D-Ribose 9. The decrease in blood glucose comes along with increases in insulin levels. The glucose‐lowering effect occurs also if meals rich in carbohydrate or fat are ingested before uptake of D-Ribose.
No studies have investigated intake of D-Ribose in infants, young children and adolescents. Whether young age may be of special concern with regard the glucose‐lowering effects of D-Ribose is not known. However, the European Food Safety Authority Panel considers that children could be particularly vulnerable to glucose‐lowering effects of d‐ribose 9.
The European Food Safety Authority notes that there is a lack of understanding of the mechanisms responsible for the short‐term decrease in blood glucose reported in the human studies, only limited data on the dose–response relationship between D-Ribose and blood glucose levels, and uncertainty about the risk of symptomatic hypoglycaemia conditions, especially in susceptible persons.
Because the decrease in glucose levels and/or the occurrence of transient symptomatic hypoglycaemia (as reported in one case) is considered adverse, based on the human studies, the lowest observed adverse effect level (LOAEL) would be at intakes of 10 g of D-Ribose. Concerning the human data, taking the above issues into account, the European Food Safety Authority concludes that 5 g per day, equivalent to 70 mg/kg body weight per day, would be the No Observed Adverse Effect Level (NOAEL) with respect to hypoglycaemia that can be considered applicable for adults. For children, the European Food Safety Authority acknowledges the lack of human data directly relevant for this population group.
Based on the No Observed Adverse Effect Level (NOAEL) of 3,600 mg/kg body weight per day derived from the subchronic toxicity study in rats, an acceptable level of intake would be up to 36 mg/kg body weight per day. This is half the NOAEL value identified in the human studies for adults with respect to hypoglycaemia. The European Food Safety Authority concludes that 36 mg/kg body weight per day would also take into account the potentially increased sensitivity of certain population groups to hypoglycaemia, including children.
The body weight therefore concludes that the D-Ribose is safe for the general population at intake levels up to 36 mg/kg body weight per day 9.
Fenstad et al. 10 found significantly higher levels of uric acid (which were, however, still in the normal range) following oral administration of 10 g D-Ribose when compared to the dose of 2 g at 30 and 60 min, no changes in serum uric acid concentrations were reported by Thompson et al. 11 following ingestion of 10 g d‐ribose and in the study of Gross et al. 12, during or after treatment with 27–89 g d‐ribose. Gastrointestinal symptoms were noted at single intakes of around 70 g 12.
The occurrence of gastrointestinal problems, short‐term hypoglycaemia and hyperuricemia in some of the studies presented and the lack of long‐term studies analyzing the effect of high doses of D-Ribose, does not allow determining the safety and tolerance of a long‐term intake of D-Ribose 9.
D-ribose health concerns
As a reducing monosaccharide, Ribose has the ability to react with proteins to produce glycated derivatives 13. Glycation with Ribose (ribosylation) gives rise to advanced glycation end products (AGEs) more rapidly than glycation with D-glucose which requires a relatively long time 14. Ribosylation (glycation with Ribose) rapidly induces alpha synuclein to form highly cytotoxic molten globules of advanced glycation end products. Alpha synuclein (α-Syn) is the main component of Lewy bodies which are associated with several neurodegenerative diseases such as Parkinson’s disease 1. D-Ribose, however, is also closely associated with many fundamental processes in cellular metabolism. Seuffer 15 has determined the concentration of D-ribose (0.01–0.1 mM) present in cerebrospinal fluid (CSF) in the brain. D-Ribose is used to synthesize nucleotides, nucleic acids, glycogen, and other important metabolic products. D-Ribose is also formed in the body from conversion of D-glucose via the pentose phosphate pathway. Thus, D-Ribose is present both intracellularly and extracellularly, and has opportunities to react with proteins and produce glycated derivatives. For this reason, glycation of Alpha synuclein protein with D-ribose needs to be investigated.
In vitro studies showed that D-Ribose induces protein misfolding rapidly leading to globular-like aggregations that are cytotoxic to neuronal cells 16. Intraperitoneal injection of D-Ribose for 30 days revealed high levels of glycated proteins and advanced glycation end products (AGEs) in the blood and brain of wild type mice. The mice also exhibited impairment of spatial learning and memory 17. More recently, D-Ribose was found to be increased in the urine of type 2 diabetic patients 18, suggesting that diabetic patients may be suffered from metabolic imbalance of not only D-Glucose but also D-Ribose 19.
Ribose intake from the diet
Ribose is reported to be present in meat in varying amounts, e.g. 1–524 mg/100 g beef meat 20 and 6.4–39.5 mg/100 g chicken meat 21, with these differences probably attributable not only to the origin of the meat but also to differences in sampling or analytical methods. Lilyblade and Peterson 22 reported that the content of ribose increases post‐mortem during storage of chicken meat. The cooking process is reported to reduce the quantity of free ribose as this is involved in Maillard reactions.
RNA and free nucleotides are reported to be degraded in the intestinal tract and free ribose may partially be released by the action of nucleosidases 23.
Due to the lack of data in the literature, the intake levels of d‐ribose from the diet are largely unknown.
Ribose inside the body production
Based on the estimated mean daily intake of carbohydrates in the UK, i.e. 252 and 198 g in men and women, respectively, 24 and assuming (i) that 2/3 of these would be composed of glucose, and (ii) that 2–9% of glucose is utilised via the pentose phosphate pathway 25, it is estimated that the daily endogenous production of d‐ribose through the pentose phosphate pathway ranges from approximately 3–15 g/day. Additional information on the extent of the daily endogenous production of d‐ribose in the literature is lacking. The information is insufficient to draw conclusions on the extent of the daily endogenous production of d‐ribose.
D-ribose dosage
Fibromyalgia and chronic fatigue syndrome patients were given 5 grams three times daily 26.
Based on the No Observed Adverse Effect Level (NOAEL) of 3,600 mg/kg body weight per day derived from the subchronic toxicity study in rats, an acceptable level of intake would be up to 36 mg/kg body weight per day. This is half the NOAEL value identified in the human studies for adults with respect to hypoglycaemia. The European Food Safety Authority concludes that 36 mg/kg body weight per day would also take into account the potentially increased sensitivity of certain population groups to hypoglycaemia, including children.
The body weight therefore concludes that the D-Ribose is safe for the general population at intake levels up to 36 mg/kg body weight per day 9.
D-ribose side effects
While oral administration of ribose is generally safe, studies also revealed that high dose ribose may induce diarrhea 27. During a post‐marketing surveillance programme the adverse reactions reported, ordered by frequency, were upset stomach (5); transient‐elevated blood sugar in diabetic customers (4); transient racing heart (3); rash (3); itching (3); diarrhoea (3); constipation (2); swollen leg (2); felt funny (1); weakness/fatigue (1); headache (1); light‐headedness (1); excess gas (1); muscle tightness (1) 9.
A study Wu et al. 28 showed that feeding D-Ribose daily through gavage to mice for 6 months is correlated with cognitive impairment. Mice treated with D-Ribose (3.75 g/kg per day) exhibited learning and memory decline and anxiety-like behavior, accompanied by Aβ-like deposition and Tau hyperphosphorylation in their brain, especially the hippocampus. These features are Alzheimer’s disease-like, both with regards to pathology (abnormal modifications and aggregations of Tau protein and Aβ peptide) and pathophysiology (memory loss accompanied by anxiety-like behavior). This suggests that D-Ribose-gavaged mouse model may be useful to study age-related cognitive impairment 29 and diabetic encephalopathy, conditions in which Aβ deposition and Tau hyperphosphorylation are found 30.
The D-Ribose-induced high level of Aβ-like deposition in the hippocampus demonstrates that mice can develop Aβ-like deposits even though the APP gene sequence of mice differs from that of humans 31. Previously, other scientists employed mice in a methanol-gavage experiment and found that methanol can distinctly promote hyperphosphorylation of Tau, but not Aβ deposition 32. However, by oral administration of D-Ribose, Aβ-like deposition can be induced under the experimental conditions. Amyloid β deposit is clear and robust.
D-Ribose is significantly increased in the brain of D-Ribose-treated mice. However, it did not markedly increase advanced glycation end levels in the brain, serum, liver and kidney under the experimental conditions. However, intraperitoneal injection of similar concentrations of D-Ribose for 30 days, as described 33, resulted in advanced glycation end accumulation in the mouse blood and brain. Whether D-Ribose elevates levels of advanced glycation ends in mice or not depends on the mode of administration. Thus, impairment of cognitive ability by oral administration of D-Ribose is probably not related to brain advanced glycation end accumulation, but rather the abnormally high levels of Aβ-like deposition and Tau hyperphosphorylation in mouse brain.
To compare the effects between D-Ribose and D-Glucose, oral administration of D-Glucose did not affect cognition. Even though D-Glucose showed elevated pS396 levels in the hippocampus and AT8 reactivity in the cortex, the elevations were not clearly seen by immunohistochemical staining of sections using antibodies AT8, pS396, p214 and T181. D-Glucose was found to reduce the percentage of open arm entries of mice in the elevated plus-maze test. However, the D-Glucose-gavaged mice did not show an abnormal behavior in the open field test, indicating that D-Glucose-gavaged mice may not be anxious. Oral administration of D-Glucose did not cause high levels of Aβ-like deposits. In contrast, gavage of D-Ribose resulted not only in high levels of Aβ-like deposition and Tau phosphorylation in the hippocampus, but also in memory loss and anxiety-like behavior.
The long-term administration of D-Ribose did not cause any significant changes in the body weight compared to mice treated with D-Glucose. Fasting blood sugar levels and advanced glycation end levels did not differ between the groups. Except for memory loss and anxiety, both D-Ribose- and D-Glucose-gavaged mice did not show additional abnormal behavior such as motor dysfunction, reduced muscle strength and impaired co-ordination.
As described by Segal and colleagues 34, D-Ribose can cause insulin release and decrease blood D-Glucose. Our results are similar to their findings. Since mice were gavaged with the sugar, it needs more insulin to regulate it. The secreted insulin induced lower level of blood sugar 35.
In Alzheimer’s disease, memory disorder is known as the most important symptom. Behavioral and psychiatric symptoms like anxiety and depression may also follow the memory loss. In this study 28, D-Ribose-treated mice exhibit an anxious symptom in open field test and elevated plus maze. The anxiety-like effect of D-Ribose on mice should be further investigated. Forced swim test showed that the mice under D-Ribose treatment behave more immobile, which indicates the depression state. While tail suspension test did not show any difference in the immobility time. Administration of D-Glucose as a control did not show anxiety-like behavior and significant depression state. Take all those results, oral administration of D-Ribose induces not only memory impairment but also anxious behavior in mice 28.
Oral administration of D-Ribose to mice also causes impaired memory functions, Tau hyperphosphorylation and Aβ-like deposition 28. These data suggest that an imbalance of D-Ribose metabolism may play a role in diabetic encephalopathy 28.
In summary, oral administration of D-Ribose leads to the impairment of cognitive ability and anxious behavior in mice 28. Meanwhile, D-Ribose-gavaged mice suffer from both Aβ-like deposition and Tau hyperphosphorylation in their brain, especially in the hippocampus 28. These abnormal modifications and the pathological aggregation of these proteins are probably correlated with the memory loss and anxiety-like behavior after D-Ribose treatment 28.
References- Chen L, Wei Y, Wang X, He R. Ribosylation Rapidly Induces α-Synuclein to Form Highly Cytotoxic Molten Globules of Advanced Glycation End Products. Petrucelli L, ed. PLoS ONE. 2010;5(2):e9052. doi:10.1371/journal.pone.0009052. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2816216/
- Heaf DJ, Davies JI. The effect of RNA supplementation of rat diets on the composition of body fluids. The British journal of nutrition. 1976;36:381–402
- PURINE NUCLEOSIDES. VI. FURTHER METHYLATION STUDIES OF NATURALLY OCCURRING PURINE NUCLEOSIDES. Biochemistry. 1964 Apr;3:494-500. https://www.ncbi.nlm.nih.gov/pubmed/14188163/
- Influence of ribose, adenosine, and “AICAR” on the rate of myocardial adenosine triphosphate synthesis during reperfusion after coronary artery occlusion in the dog. Mauser M, Hoffmeister HM, Nienaber C, Schaper W. Circ Res. 1985 Feb; 56(2):220-30.
- Biosynthesis of riboflavin: mechanism of formation of the ribitylamino linkage. Keller PJ, Le Van Q, Kim SU, Bown DH, Chen HC, Kohnle A, Bacher A, Floss HG. Biochemistry. 1988 Feb 23; 27(4):1117-20.
- MacCarter D, Vijay N, Washam M, Shecterle L, Sierminski H, St Cyr JA. D-ribose aids advanced ischemic heart failure patients. International journal of cardiology. 2009;137:79–80 https://www.ncbi.nlm.nih.gov/pubmed/18674831
- Teitelbaum JE, Johnson C, St Cyr J. The use of D-ribose in chronic fatigue syndrome and fibromyalgia: a pilot study. J Altern Complement Med. 2006;12:857–862. https://www.ncbi.nlm.nih.gov/pubmed/17109576
- Perlmutter NS, Wilson RA, Angello DA, Palac RT, Lin J, Brown BG. Ribose facilitates thallium-201 redistribution in patients with coronary artery disease. J Nucl Med. 1991;32:193–200. http://jnm.snmjournals.org/content/32/2/193.long
- Safety of d‐ribose as a novel food pursuant to Regulation (EU) 2015/2283. EFSA Journal 31 May 2018 https://doi.org/10.2903/j.efsa.2018.5265
- Fenstad ER, Seifert JG, Gazal O and St Cyr JA, 2000. D‐Ribose’s Effect on Carbohydrates and Purine Metabolism. Human Performance Laboratory, St. Cloud State University (MN)/Bioenergy, Inc., Minneapolis. Unpublished report.
- Thompson J, Neutel J, Homer K, Tempero K, Shah A and Khankari R, 2014. Evaluation of D‐ribose pharmacokinetics, dose proportionality, food effect, and pharmacodynamics after oral solution administration in healthy male and female subjects. Journal of Clinical Pharmacology, 54, 546–554.
- Gross M, Reiter S and Zöllner N, 1989. Metabolism of D‐ribose administered continuously to healthy persons and to patients with myoadenylate deaminase deficiency. Klinische Wochenschrift, 67, 1205–1213.
- Han C, Lu Y, Wei Y, Liu Y, He R. D-Ribose Induces Cellular Protein Glycation and Impairs Mouse Spatial Cognition. Li X-J, ed. PLoS ONE. 2011;6(9):e24623. doi:10.1371/journal.pone.0024623. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3169629/
- Nonenzymatic browning in vivo: possible process for aging of long-lived proteins. Monnier VM, Cerami A. Science. 1981 Jan 30; 211(4481):491-3.
- Seuffer R. [A new method for the determination of sugars in cerebrospinal fluid (author’s transl)]. J Clin Chem Clin Biochem. 1977;15:663–668. https://www.ncbi.nlm.nih.gov/pubmed/604418
- Chen L, Wei Y, Wang XQ, He RQ. Ribosylation Rapidly Induces a-Synuclein to Form Highly Cytotoxic Molten Globules of Advanced Glycation End Products. PLoS ONE. 2010;5:e9052. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2816216/
- Han C, Lu Y, Wei Y, Liu Y, He RQ. D-ribose induces cellular protein glycation and impairs mouse spatial cognition. PLoS ONE. 2011;6:e24623 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3169629/
- Su T, He RQ. The Abnormally High Level of Uric D-Ribose for Type-2 Diabetics. Prog Biochem Biophys. 2013;40:816–825.
- Su T, He RQ. D-ribose, an overlooked player in type 2 diabetes mellitus? Sci China Life Sci. 2014;57:361 https://www.ncbi.nlm.nih.gov/pubmed/24448907
- Gazzani G and Cuzzoni M, 1985. Sulla caratterizzazione della carne bovina proveniente da vitelli e da animali adulti. Rivista della Societa Italiana di Scienze Alimentari, 14, 369–372.
- Aliani M and Farmer LJ, 2002. A postcolumn derivatization method for determination of reducing and phosphorylated sugars in chicken by high performance liquid chromatography. Journal of Agricultural and Food Chemistry, 50, 2760–2766.
- Lilyblade AL and Peterson DW, 1962. Inositol and free sugars in chicken muscle post‐mortem. Journal of Food Science, 245–249.
- Carver JD and Walker WA, 1995. The role of nucleotides in human nutrition. Nutritional Biochemistry, 6, 58–72.
- NatCen Social Research, Medical Research Council and University of Newcastle upon Tyne, 2011. Diet and Nutrition Survey of Infants and Young Children, 2011 [computer file]. 2nd Edition. Colchester, Essex: UK Data Archive [distributor], October 2013. SN: 7263, https://doi.org/10.5255/ukda-sn-7263-2
- Delgado TC, Castro MM, Geraldes CF and Jones JG, 2004. Quantitation of erythrocyte pentose pathway flux with [2–13C]glucose and 1H NMR analysis of the lactate methyl signal. Magnetic Resonance in Medicine, 51, 1283–1286.
- The Use of D-Ribose in Chronic Fatigue Syndrome and Fibromyalgia: A Pilot Study. The Journal of Alternative and Complementary Medicine VOL. 12, NO. 9. https://doi.org/10.1089/acm.2006.12.857
- Gross M, Reiter S, Zollner N. Metabolism of D-ribose administered continuously to healthy persons and to patients with myoadenylate deaminase deficiency. Klinische Wochenschrift. 1989;67:1205–13.
- Wu B, Wei Y, Wang Y, et al. Gavage of D-Ribose induces Aβ-like deposits, Tau hyperphosphorylation as well as memory loss and anxiety-like behavior in mice. Oncotarget. 2015;6(33):34128-34142. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4741441/
- Ahmed N, Ahmed U, Thornalley PJ, Hager K, Fleischer G, Munch G. Protein glycation, oxidation and nitration adduct residues and free adducts of cerebrospinal fluid in Alzheimer’s disease and link to cognitive impairment. J Neurochem. 2005;92:255–263
- Norelle L. Daly, Ralf Hoffmann, Laszlo Otvos J, Craik DJ. Role of Phosphorylation in the Conformation of Tau Peptides Implicated in Alzheimer’s Disease. Biochemistry. 2000;39:9039–9046
- Maloney B, Ge YW, Greig N, Lahiri DK. Presence of a “CAGA box” in the APP gene unique to amyloid plaque-forming species and absent in all APLP-1/2 genes: implications in Alzheimer’s disease. FASEB J. 2004;18:1288–1290
- Yang M, Lu J, Miao J, Rizak J, Yang J, Zhai R, Zhou J, Qu J, Wang J, Yang S, Ma Y, Hu X, He RQ. Alzheimer’s disease and methanol toxicity (part 1): chronic methanol feeding led to memory impairments and tau hyperphosphorylation in mice. J Alzheimers Dis. 2014;41:1117–1129.
- Han C, Lu Y, Wei Y, Liu Y, He RQ. D-ribose induces cellular protein glycation and impairs mouse spatial cognition. PLoS ONE. 2011;6:e24623
- Segal S, Foley J. The metabolism of D-ribose in man. J Clin Invest. 1958;37:719–735.
- Segal S, Foley J. The metabolism of D-ribose in man. J Clin Invest. 1958;37:719–735



{kind=link}