{kind=link}
Human eye
The human eye, the organ containing visual receptors, provides vision, with the assistance of accessory organs. These accessory organs include the eyelids and lacrimal apparatus, which protect the eye, and a set of extrinsic muscles, which move the eye.
The eye, lacrimal gland, and associated extrinsic muscles are housed in the orbital cavity, or orbit, of the skull. Each orbit is lined with the periosteum of various bones, and also contains fat, blood vessels, nerves, and connective tissues. Each eyelid, both upper and lower, has four layers—skin, muscle, connective tissue, and conjunctiva. The skin of the eyelid, which is the thinnest skin of the body, covers the lid’s outer surface and fuses with its inner lining near the margin of the lid. The eyelids are moved by the orbicularis oculi muscle (Figure 4), which acts as a sphincter and closes the lids when it contracts, and by the levator palpebrae superioris muscle, which raises the upper lid and thus helps open the eye.
The conjunctiva is a mucous membrane that lines the inner surfaces of the eyelids (palpebral conjunctiva) and folds back to cover the anterior surface of the eyeball (bulbar conjunctiva), except for its central portion (cornea).
The lacrimal apparatus consists of the lacrimal gland, which secretes tears, and a series of ducts that carry tears into the nasal cavity (Figure 3). The lacrimal gland is located in the orbit and secretes tears continuously through tiny tubules. The tears flow downward and medially across the eye. Two small ducts (the superior and inferior canaliculi) collect tears, which then flow into the lacrimal sac, located in a deep groove of the lacrimal bone. From there the tears flow into the nasolacrimal duct, which empties into the nasal cavity. Secretion by the lacrimal gland moistens and lubricates the surface of the eye and the lining of the lids. Tears also have an enzyme (lysozyme) that kills bacteria, reducing the risk of eye infections.
The extrinsic muscles of the eye arise from the bones of the orbit and insert by broad tendons on the eye’s tough outer surface. Six extrinsic muscles move the eye in different directions (Figure 5). Any given eye movement may utilize more than one extrinsic muscle, but each muscle is associated with a primary action. Figure 5 illustrates the locations of these extrinsic muscles and Table 1 lists their functions, as well as the functions of the eyelid muscles.
Figure 1. Human eye
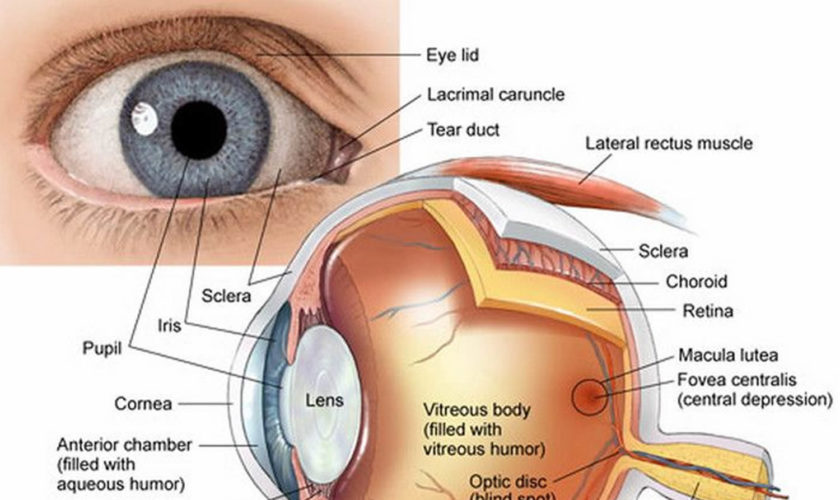
Figure 2. Eye anatomy
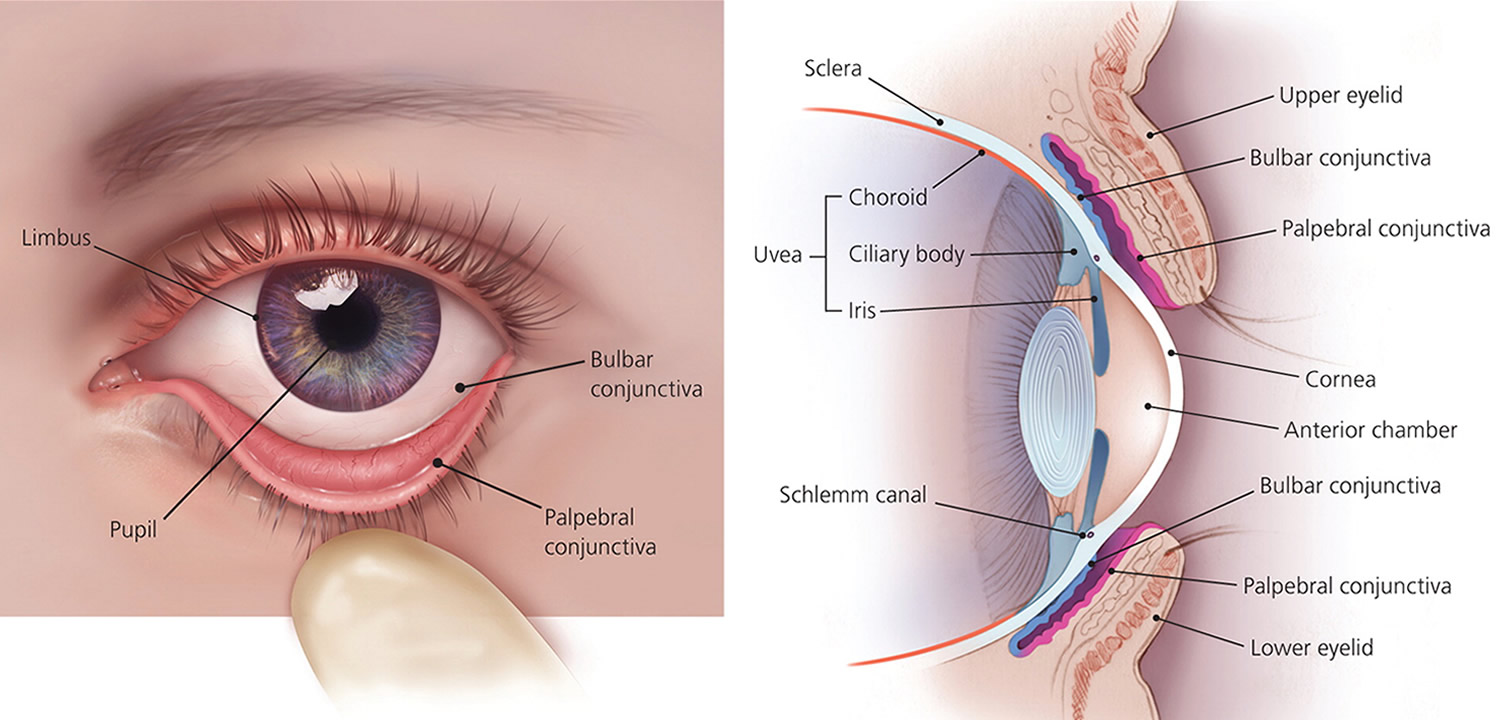
Figure 3. Lacrimal apparatus
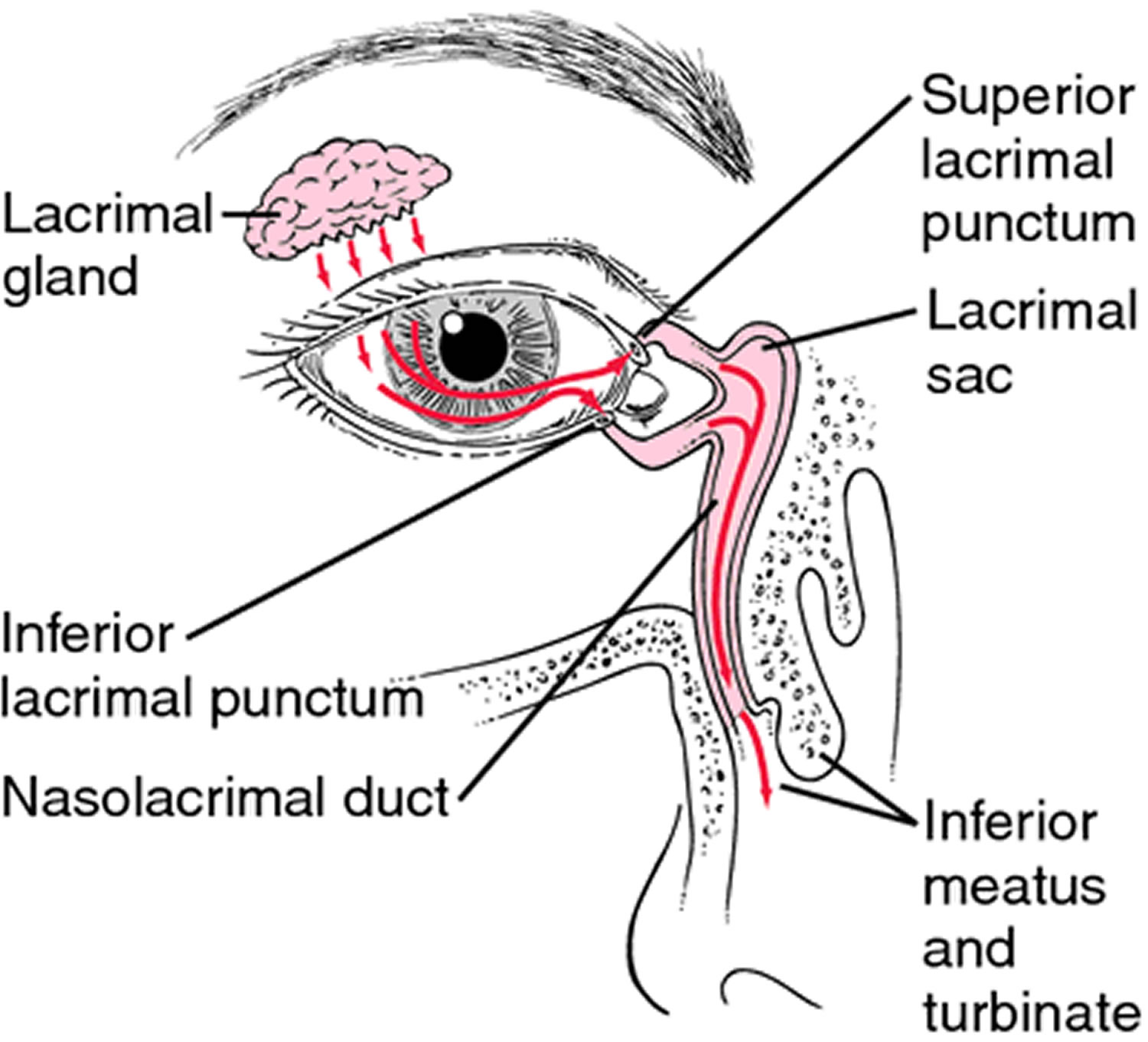
Figure 4. Orbicularis oculi muscle
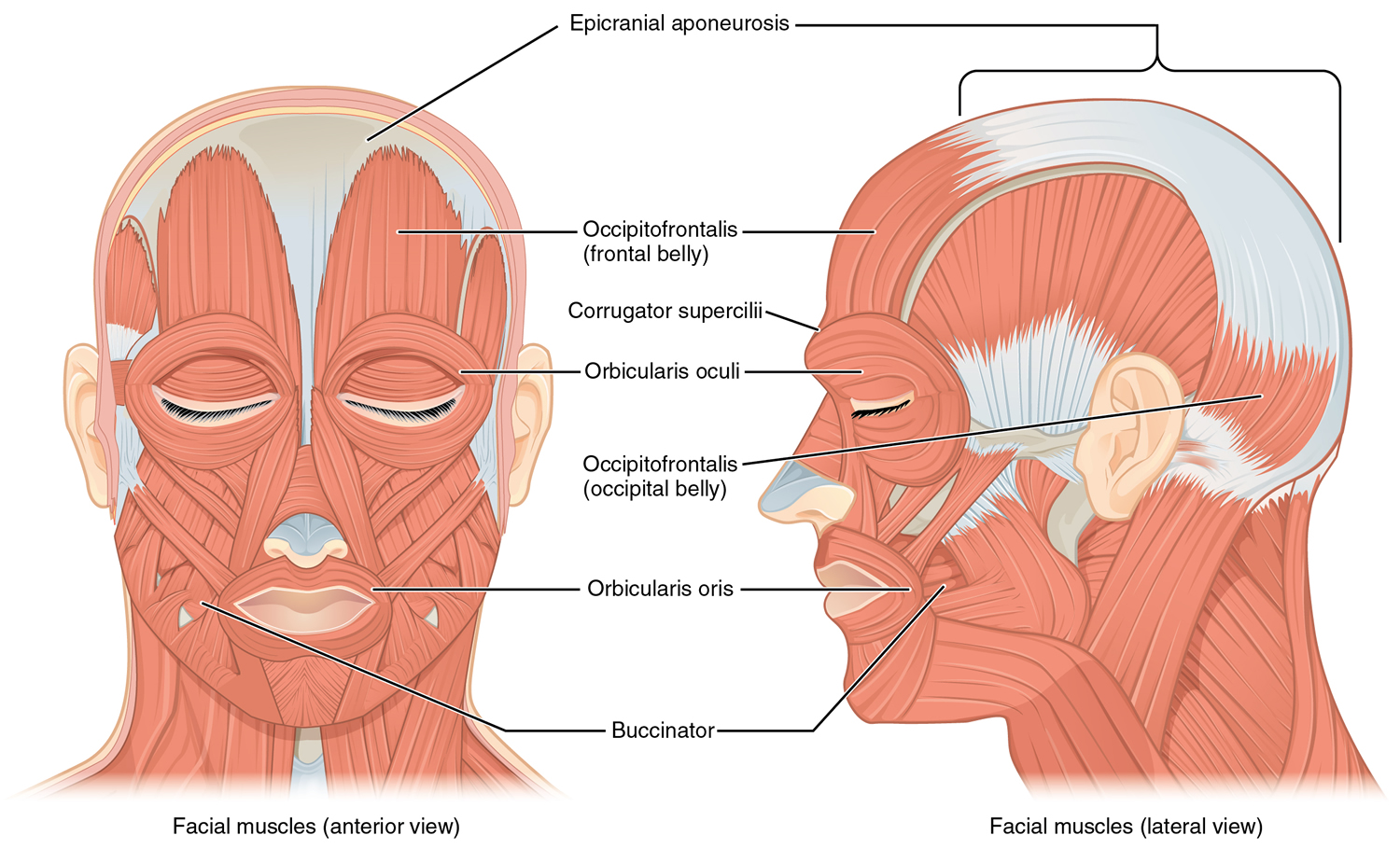
Figure 5. Eye muscles

Table 1. Muscles Associated with the Eyelids and Eyes
| Name | Innervation | Function |
| Muscles of the Eyelids | ||
| Orbicularis oculi | Facial nerve (VII) | Closes eye |
| Levator palpebrae superioris | Oculomotor nerve (III) | Opens eye |
| Extrinsic Muscles of the Eyes | ||
| Superior rectus | Oculomotor nerve (III) | Rotates eye upward and toward midline |
| Inferior rectus | Oculomotor nerve (III) | Rotates eye downward and toward midline |
| Medial rectus | Oculomotor nerve (III) | Rotates eye toward midline |
| Lateral rectus | Abducens nerve (VI) | Rotates eye away from midline |
| Superior oblique | Trochlear nerve (IV) | Rotates eye downward and away from midline |
| Inferior oblique | Oculomotor nerve (III) | Rotates eye upward and away from midline |
Structure of the Human Eye
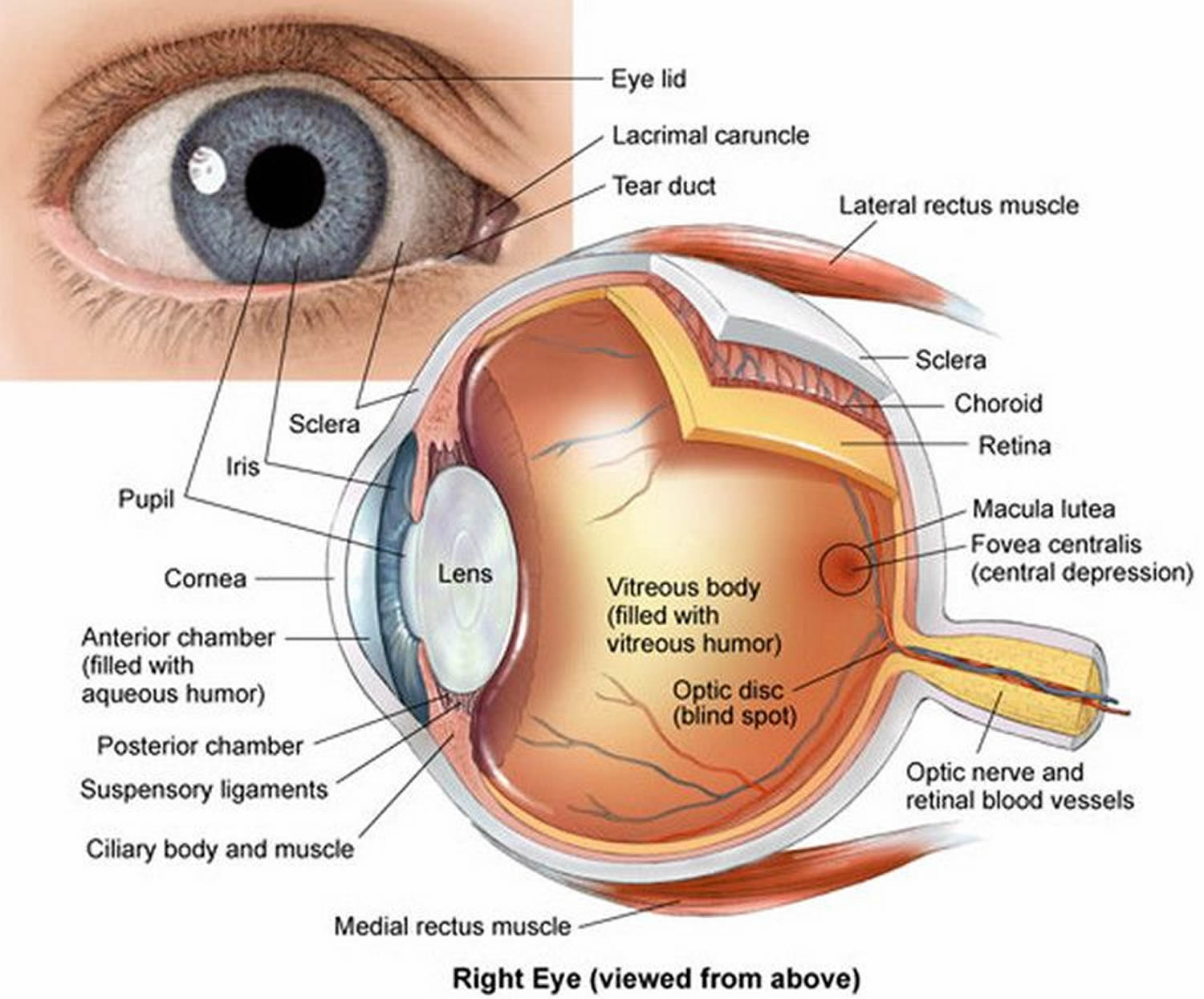
The eye is a hollow, spherical structure about 2.5 centimeters in diameter. Its wall has three distinct layers—an outer (fibrous) layer, a middle (vascular) layer, and an inner (nervous) layer. The spaces within the eye are filled with fluids that help maintain its shape.
Figure 6. Structure of the human eye
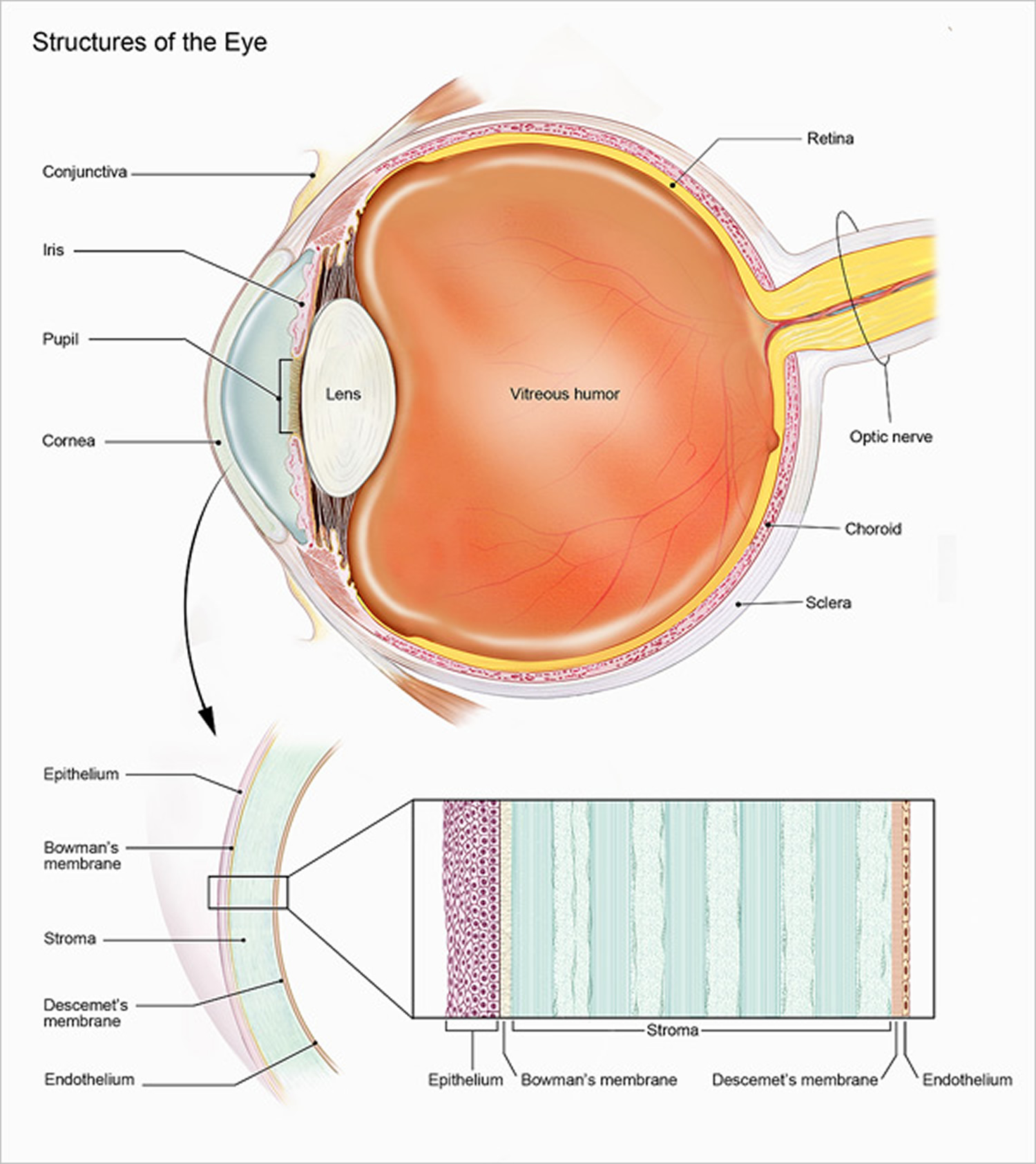
Outer Layer of the Eye
Light is often described as moving in straight lines called rays of light. The anterior sixth of the outer layer bulges forward as the transparent cornea, the window of the eye. The cornea helps focus entering light rays. The cornea is composed largely of connective tissue with a thin surface layer of epithelium. It is transparent because it contains few cells and no blood vessels, and the cells and collagenous fibers form unusually regular patterns.
The cornea is continuous with the sclera, the white portion of the eye. The sclera makes up the posterior five-sixths of the outer layer of the wall of the eye. It is opaque due to many large, disorganized, collagenous and elastic fibers. The sclera protects the eye and serves as an attachment for the extrinsic muscles. In the back of the eye, the optic nerve and certain blood vessels pierce the sclera.
Middle Layer of the Eye
The middle layer of the wall of the eye includes the choroid coat, ciliary body, and iris. The choroid coat, in the posterior five-sixths of the globe of the eye, is loosely joined to the sclera and is honeycombed with blood vessels, which nourish surrounding tissues. The choroid coat also has many pigment-producing melanocytes. The melanin that these cells produce absorbs excess light, which helps keep the inside of the eye dark.
The ciliary body is the thickest part of the middle layer of the wall of the eye. It extends forward and inward from the choroid coat and forms a ring inside the front of the eye. Within the ciliary body are many radiating folds called ciliary processes and groups of smooth muscle cells that constitute the ciliary muscle.
Many strong but delicate fibers, called suspensory ligaments, extend inward from the ciliary processes and hold the transparent lens in position. The distal ends of these fibers attach along the margin of a thin capsule that surrounds the lens. The body of the lens lies directly behind the iris and pupil and is composed of highly specialized epithelial cells called lens fibers. The cytoplasm of these cells is the transparent substance of the lens.
The ciliary muscle and suspensory ligaments, along with the structure of the lens itself, enable the lens to adjust shape to facilitate focusing, a phenomenon called accommodation. The lens is enclosed by a clear capsule composed largely of elastic fibers. This elastic nature keeps the lens under constant tension, and enables it to assume a globular shape. The suspensory ligaments attached to the margin of the capsule are also under tension. When they pull outward, flattening the capsule and the lens inside, the lens focuses on distant objects. However, if the tension on the suspensory ligaments relaxes, the elastic lens capsule rebounds, and the lens surface becomes more convex—focused for viewing closer objects.
The ciliary muscle controls the actions of the suspensory ligaments in accommodation. One set of ciliary muscle cells extends back from fixed points in the sclera to the choroid coat. When the muscle contracts, the choroid coat is pulled forward and the ciliary body shortens. This action relaxes the suspensory ligaments, and the lens thickens in response. When the ciliary muscle relaxes, tension on the suspensory ligaments increases, and the lens becomes thinner and less convex again.
More than 90% of the proteins in a lens cell are lens crystallins, which aggregate into fibers. These proteins, along with the absence of organelles that scatter light (mitochondria, endoplasmic reticula, and nuclei), account for the transparency of the lens. An eye disorder common in older people is cataract. The lens or its capsule slowly becomes cloudy and opaque. Cataracts are treated on an outpatient basis with surgery to remove the clouded lens and replace it with an artificial lens. Without treatment, cataracts eventually cause blindness.
The iris is a thin diaphragm composed mostly of connective tissue and smooth muscle fibers. From the outside, the iris is the colored part of the eye. The iris extends forward from the periphery of the ciliary body and lies between the cornea and lens. The iris divides the space (anterior cavity) separating these parts into an anterior chamber (between the cornea and the iris) and a posterior chamber containing the lens (between the iris and the vitreous body).
The epithelium on the inner surface of the ciliary body secretes a watery fluid called aqueous humor into the posterior chamber. The fluid circulates from this chamber through the pupil, a circular opening in the center of the iris, and into the anterior chamber. Aqueous humor fills the space between the cornea and lens, helps nourish these parts, and aids in maintaining the shape of the front of the eye. Aqueous humor leaves the anterior chamber through veins and a special drainage canal, the scleral venous sinus (canal of Schlemm). This sinus is in the wall of the anterior chamber at the junction of the cornea and the sclera.
The iris controls the size of the pupil, through which light passes as it enters the eye. The contractile cells of the iris are organized into two groups, a circular set and a radial set. The circular set (pupillary constrictor) is smooth muscle and acts as a sphincter. When the muscle cells contract, the pupil gets smaller, and less light enters. Bright light stimulates the circular muscles to contract, which decreases the amount of light entering the eye. The radial set (pupillary dilator) is composed of specialized contractile epithelial cells (myoepithelial cells). When these cells contract, the pupil’s diameter increases and more light enters. Dim light stimulates the radial cells to contract, which increases the amount of light entering the eye.
Inner Layer of the Eye
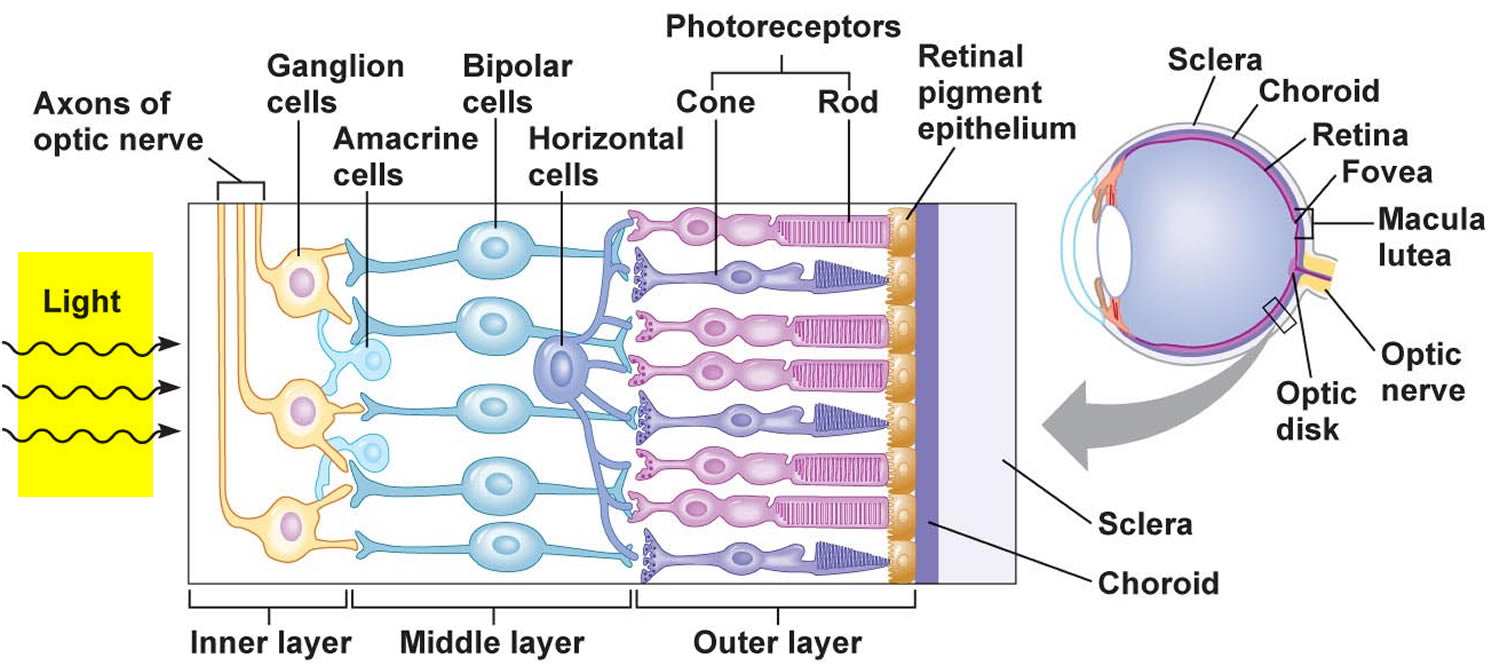
The inner layer of the wall of the eye consists of the retina, which contains the visual receptor cells (photoreceptors). The retina is a nearly transparent sheet of tissue continuous with the optic nerve in the back of the eye and extending forward as the inner lining of the eyeball. The retina ends just behind the margin of the ciliary body.
The retina is thin and delicate, but its structure is quite complex. It has a number of distinct layers, as Figure 7 illustrates. In the central region of the retina is a yellowish spot called the macula lutea. A depression in its center, called the fovea centralis, is in the region of the retina that produces the sharpest vision (see Figure 6).
Just medial to the fovea centralis is an area called the optic disc. Here, nerve fibers from the retina leave the eye and form the optic nerve. Because the optic disc region does not have photoreceptors, it is commonly known as the blind spot of the eye. A central artery and vein also pass through the optic disc. These vessels are continuous with the capillary networks of the retina. Along with vessels in the underlying choroid coat, they supply blood to the cells of the inner layer.
The space bounded by the lens, ciliary body, and retina is the largest compartment of the eye and is called the posterior cavity. It is filled with a transparent, jellylike fluid called vitreous humor, which along with collagen fibers forms the vitreous body. The vitreous body supports the internal parts of the eye and helps maintain the eye’s shape.
Photoreceptors are modified neurons of two distinct kinds, as Figure 7 illustrates. One group of photoreceptors, called rods, have long, thin projections at their ends, and provide vision without color, in shades of gray. The other group of photoreceptors, called cones, have short, blunt projections, and provide color vision. There is only one type of rod, but there are three types of cones. A human eye has 125 million rods and 7 million cones. Rods and cones are in a deep part of the retina. They are closely associated with an adjacent layer of retinal pigment epithelium (RPE) that absorbs light the photoreceptors do not absorb. With the pigment of the choroid coat, the retinal pigment epithelium keeps light from reflecting off surfaces inside the eye.
Photoreceptors are stimulated only when light reaches them. A light image focused on an area of the retina stimulates some photoreceptors, which results in impulses traveling to the brain. However, the impulses leaving in response to each activated photoreceptor deliver only a fragment of the information required for the brain to interpret a complete scene.
Rods and cones contribute to different aspects of vision. Rods are hundreds of times more sensitive to light than cones and therefore can provide vision in dim light, but without color. Cones detect color but are less sensitive and do not respond in dim light.
Rods and cones also differ in the sharpness of the perceived images, or visual acuity. Cones provide sharp images, and rods provide more general outlines of objects. Rods give less precise images because axon branches from rods undergo convergence. Because of this, impulses from a group of rods are conducted to the brain on a single nerve fiber. Thus, if a point of light stimulates a rod, the brain cannot tell which one of many receptors has been stimulated. Convergence of impulses is less common among cones. When a cone is stimulated, the brain can pinpoint the stimulation more accurately.
The fovea centralis, the area of sharpest vision, does not have rods but contains densely packed cones with few or no converging fibers. Also in the fovea centralis, the overlying layers of the retina and the retinal blood vessels are displaced to the sides, more fully exposing photoreceptors to incoming light. Consequently, to view something in detail, a person moves the eyes so that the important part of an image falls on the fovea centralis.
Both rods and cones contain light-sensitive pigments that decompose when they absorb light energy. The light-sensitive biochemical in rods is called rhodopsin or visual purple. In the presence of light, rhodopsin molecules are broken down into a colorless protein called opsin and a yellowish substance called retinal (retinene) that is synthesized from vitamin A.
Decomposition of rhodopsin molecules activates an enzyme that initiates a series of reactions altering the permeability of the rod cell membrane. As a result, a complex pattern of nervous stimulation originates in the retina. The impulses are conducted away from the retina along the optic nerve into the brain, where they are interpreted as vision. In bright light, nearly all of the rhodopsin in the rods of the retina decomposes, greatly reducing rod sensitivity. In dim light, however, regeneration of rhodopsin from opsin and retinal is faster than rhodopsin breakdown. ATP provides the energy required for this regeneration.
The light-sensitive pigments in cones, as in the rods, are composed of retinal and protein. In cones, however, three types of opsin proteins, different from the protein portion of rhodopsin found in rods, combine with retinal to form the three cone pigments. The three types of cones each contain one of these three photopigments.
Poor vision in dim light, called nightblindness, results from vitamin A deficiency. Lack of the vitamin reduces the supply of retinal, rhodopsin production falls, and rod sensitivity is low. Nightblindness is treated by supplementing the diet with vitamin A.
The wavelength of light determines the color that the brain perceives from it. For example, the shortest wavelengths of visible light are perceived as violet, and the longest are perceived as red. One type of cone pigment (erythrolabe) is most sensitive to red light, another (chlorolabe) to green light, and the third (cyanolabe) to blue light. The color a person perceives depends on which cones the light in a given image stimulates. If all three sets of cones are stimulated with equal intensity, the person senses the light as white, and if none are stimulated, the person senses black. Different forms of colorblindness result from the body’s inability to produce different types of cone pigments.
Figure 7. The retina (inside human eye)
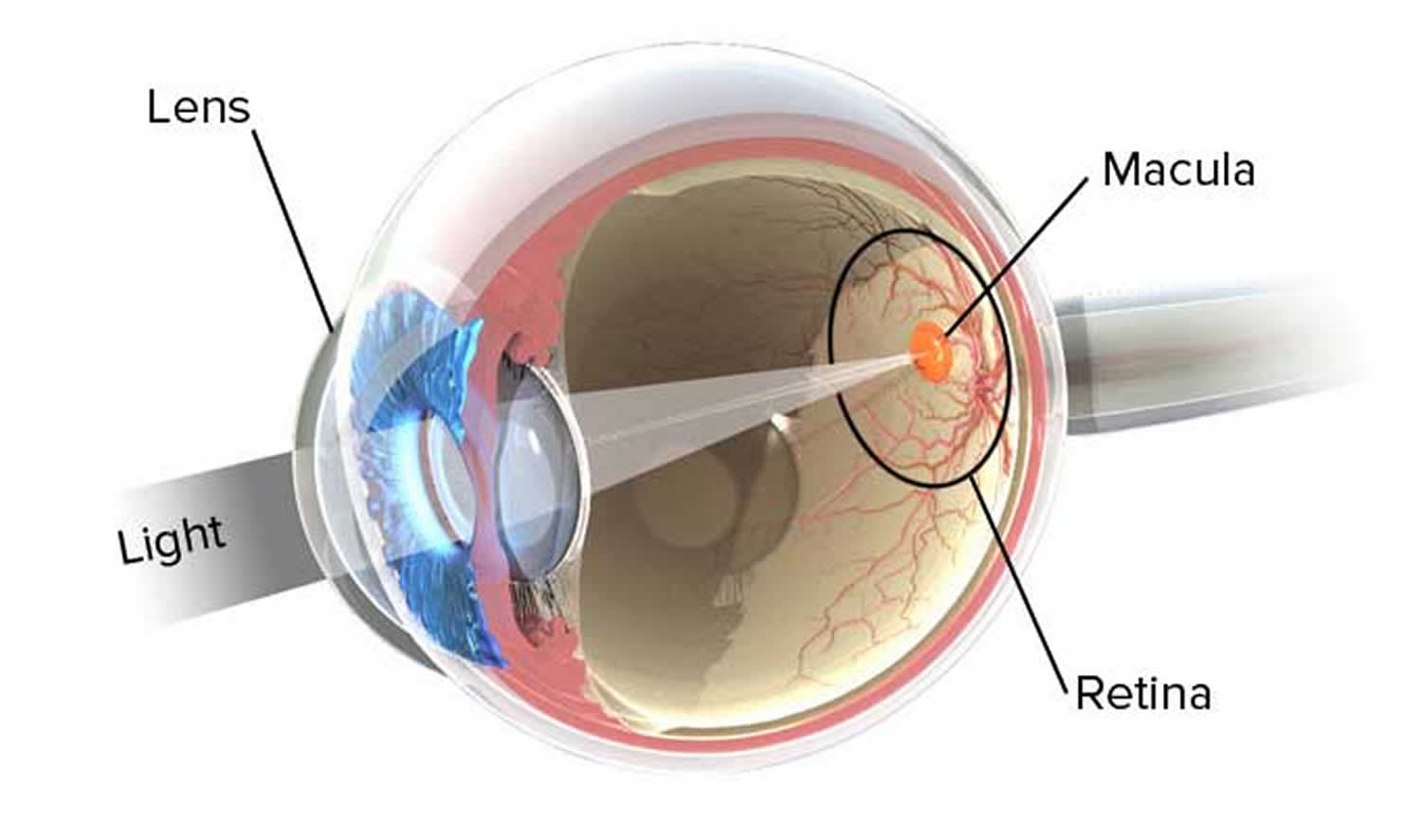
When a person sees an object, either the object is giving off light or it is reflecting light from another source. Rays of light enter the eye, and an image of the object is focused on the retina. Focusing bends the light rays, a phenomenon called refraction, so the image falls on the fovea centralis (Figure 8). Refraction occurs when light rays pass at an oblique angle from a substance of one optical density into a substance of a different optical density. This happens at the curved surface between the air and the cornea and at the curved surfaces of the lens itself. A lens with a convex surface (as in the eye) causes light rays to converge.
The convex surface of the cornea refracts light rays from outside objects. The convex surface of the lens and, to a lesser extent, the surfaces of the fluids in the chambers of the eye then refract the light again. If eye shape is normal, light rays focus sharply on the retina, much as a motion picture image is focused on a screen for viewing. Unlike the motion picture image, however, the image that forms on the retina is upside down and reversed from left to right. The visual cortex interprets the image in its proper position.
Figure 8. Normal eye refraction (perfect image on the Macula’s Fovea)
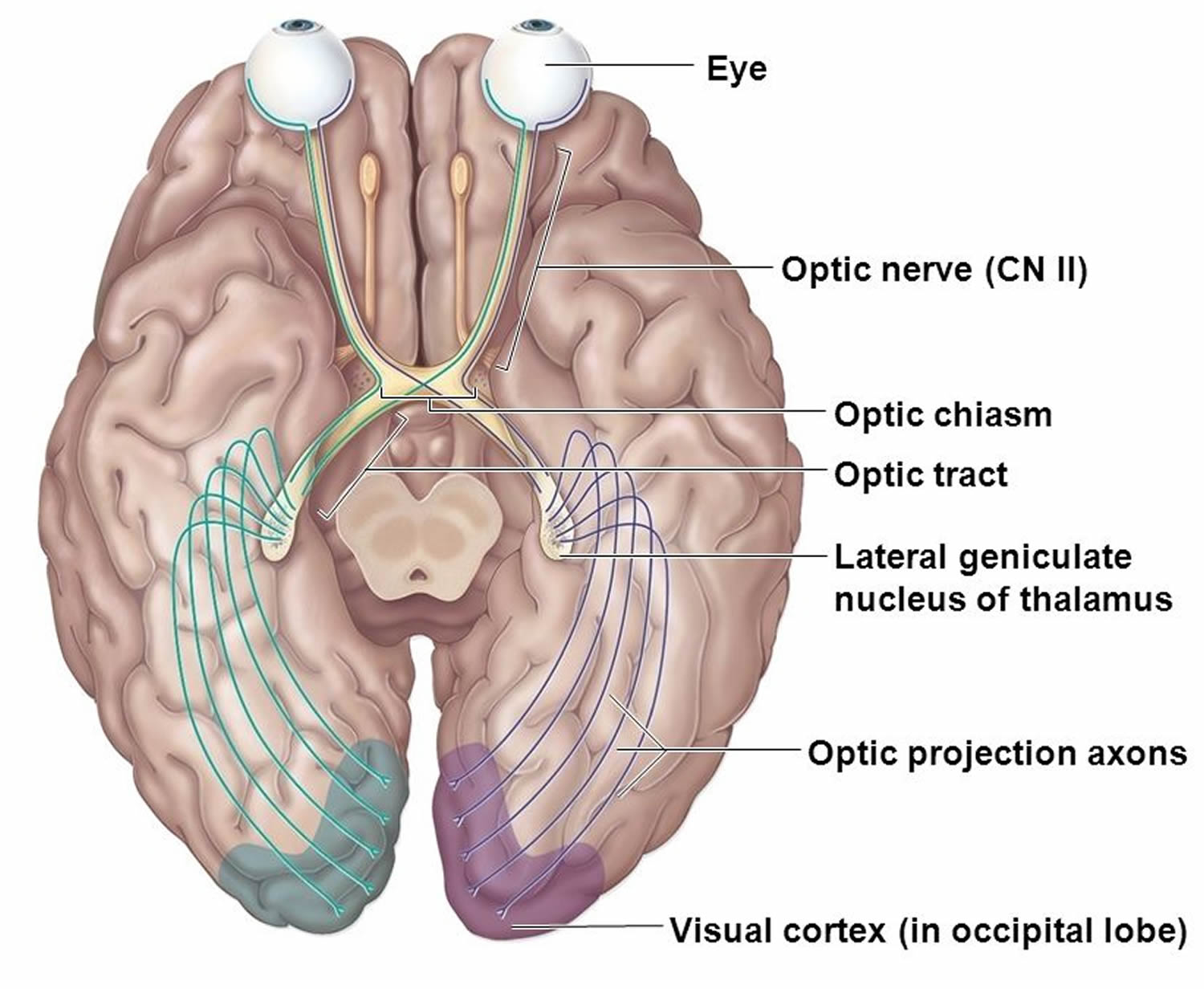
Visual Pathways
Visual pathways conduct impulses from the retina to the visual cortex, where they are perceived as vision (see Figure 9). The pathways begin as the axons of the retinal neurons leave the eyes to form the optic nerves. Just anterior to the pituitary gland, these nerves give rise to the X-shaped optic chiasma. Within the chiasma, some of the fibers cross over. More specifically, the fibers from the nasal (medial) half of each retina cross over, but those from the temporal (lateral) sides do not. Thus, fibers from the nasal half of the left eye and the temporal half of the right eye form the right optic tract, and fibers from the nasal half of the right eye and the temporal half of the left eye form the left optic tract.
Just before the nerve fibers reach the thalamus, a few of them enter nuclei that function in various visual reflexes. Most of the fibers, however, enter the thalamus and synapse in its posterior portion (lateral geniculate body). From this region, the visual impulses enter nerve pathways called optic radiations, which lead to the visual cortex of the occipital lobes.
Figure 9. Visual pathway