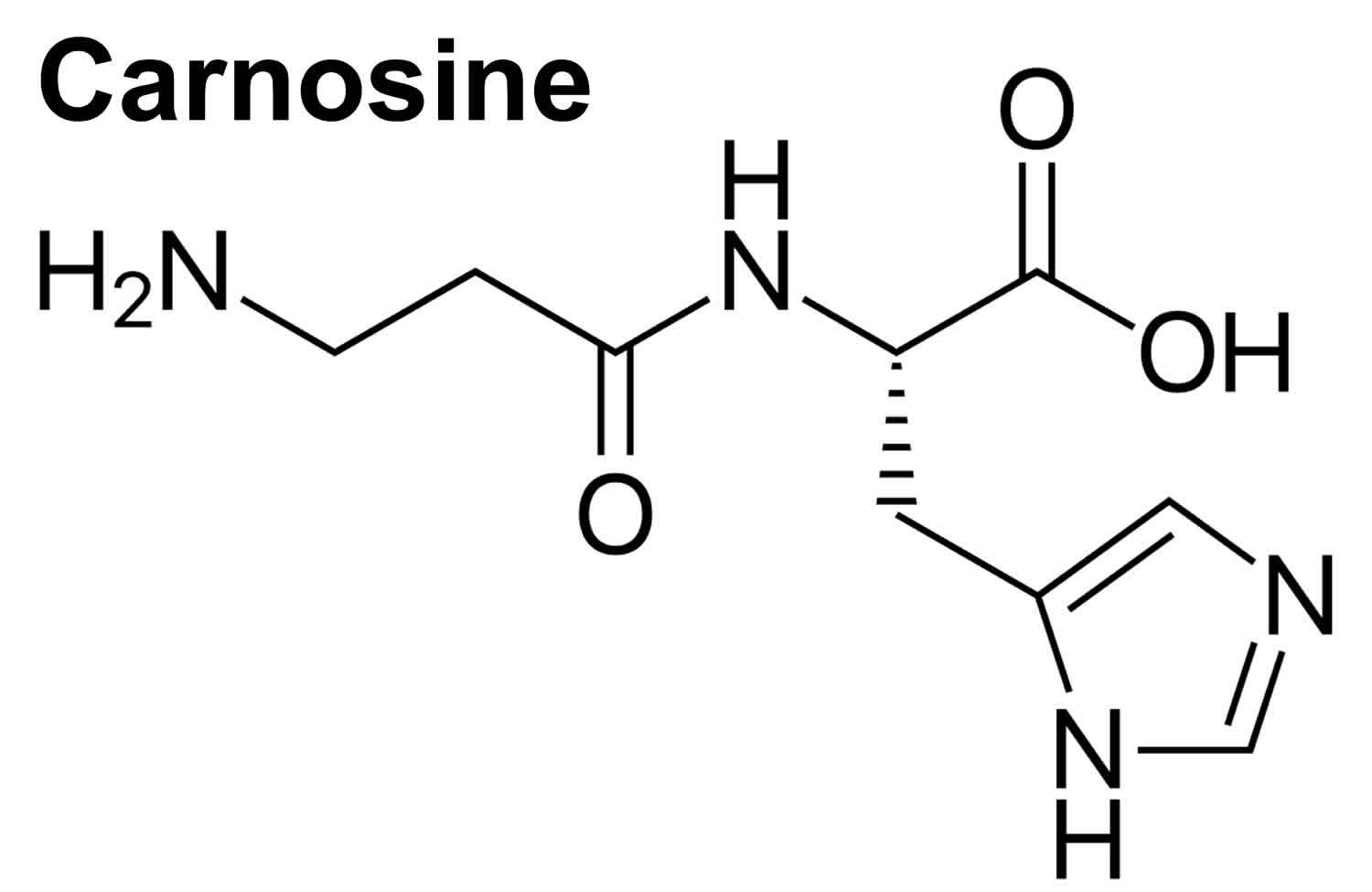
What is carnosine
Carnosine (beta-alanyl-L-histidine) is a naturally occurring dipeptide neuropeptide (dipeptide of the amino acids beta-histidine and 3-methyl-L-histidine) found in mammalian tissues 1 and exists at particularly high concentrations (millimolar order) in the brain as well as in skeletal and cardiac muscles (up to 20 mM) 2. In the human body, concentrations of carnosine are highest in muscle tissue 3. Carnosine is one of the most abundant small-molecule compounds in skeletal muscle, with concentrations similar to those of creatine and adenosine triphosphate (ATP). Carnosine contributes to the physicochemical buffering of lactate caused by exercise in skeletal muscles and has anti-fatigue effects. Moreover, brain tissue also has fairly high concentrations of carnosine 3. Carnosine (beta-alanyl-L-histidine) is found exclusively in animal muscles (red meat) 4. Carnosine is a biomarker for the consumption of meat.
Carnosine has various advantageous characteristics, such as anti-glycation, anti-stress, and antioxidant properties, hydroxyl radical scavenging, maintenance of pH-balance, and chelation of metals including divalent zinc ion (Zn2+) and bivalent copper ion (Cu2+) 5.
Carnosine is found to be associated with serum carnosinase deficiency (carnosinemia), which is an inborn error of metabolism. Murphey and co-workers 6 reported carnosine accumulation in the serum (20-30 μmol/mL) and urine (60-200 μmol/24 h) of patients affected by a rare inherited metabolic disorder – serum carnosinase deficiency. Serum carnosinase deficiency is biochemically characterized by carnosine accumulation in the serum and urine of affected patients due to low activity of serum carnosinase 6. On the other hand, serum carnosine concentration in healthy subjects under fasting is undetectable 7. The main clinical manifestations assigned to serum carnosinase deficiency include intellectual deficiency, growth delay, seizures, twitching, lethargy, generalized muscle wasting 6, and hypotonia 8. The onset of the symptoms occurred during infancy and children presented progressive mental retardation. Serum carnosinase deficiency is an autosomal-recessive inherited metabolic disease with an estimated prevalence of 1:500,000 newborns 9. The serum carnosinase gene (CNDP1) is located in the region 18q22.3 10. To date, the most of the serum carnosinase deficiency patients reported were children with neurological dysfunction 11. Homocarnosinosis (EC 236130), which was considered as a distinct disorder, may be a severe serum carnosinase deficiency, since the presence of the enzyme homocarnosinase was not shown in human tissues 12. The phenotype of serum carnosinase deficiency patients is very variable and it was suggested that the enzyme deficiency is not pivotal to the neurological dysfunction 11.
Figure 1. Carnosine metabolic pathways
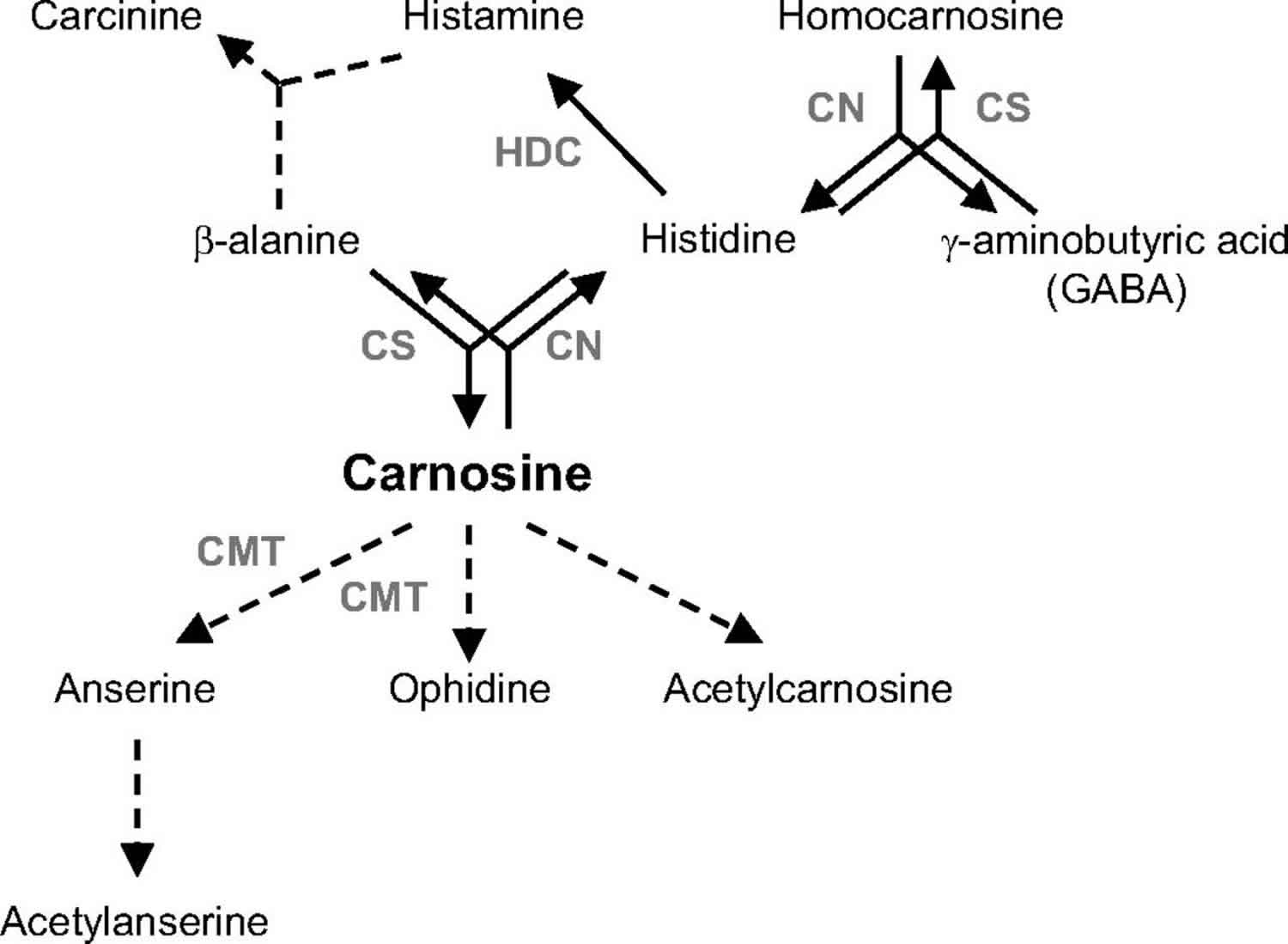
Footnote: The dipeptides carnosine and homocarnosine are synthesized by carnosine synthase (CS) and degraded by carnosinase (CN). Solid lines represent well-described reactions occurring in humans. Dashed lines represent reactions that either do not occur in humans or are poorly understood at present.
Abbreviations: HDC = l-histidine decarboxylase; CMT = carnosine-N-methyltransferase.
[Source 13 ]Carnosine benefits
Carnosine has the potential to suppress many of the biochemical changes that accompany ageing (e. g. protein oxidation, glycation, advanced glycosylation end‐product formation, and cross-linking) and associated pathologies 4. The brain may be in particular need of the antiglycosylation and antioxidant effects that carnosine provides. Glycosylation, oxidative stress, inflammation secondary to the oxidative stress, and protein cross-linking from methylglyoxal, a product of uncontrolled glucose oxidation, are all thought to contribute to the progression of Alzheimer’s Disease. There is evidence from animal studies that carnosine can reduce the toxic effects of beta-amyloid protein. Beta-amyloid (Aβ) is the abnormal protein that builds up in the brains of people with Alzheimer’s Disease and damages the surrounding brain tissue. For these reasons, increasing levels of carnosine in the brain may have a protective effect from Alzheimer’s Disease. Carnosine also exhibits some antioxidant effects. The antioxidant mechanism of carnosine is attributed to its chelating effect against metal ions, superoxide dismutase (SOD)-like activity, and ROS and free radicals scavenging ability 14.
Carnosine has been shown to be neuroprotective through different mechanisms: the prevention of oxidative stress 15, reduction of intraneuronal amyloid-βeta (Aβ) accumulation, mitochondrial dysfunctions and cognitive deficits in 3xTg-Alzheimer’s disease mice 16, as well as the inhibition of amyloid-beta (Aβ) aggregation 17, modulation of macrophage nitric oxide (NO) production and pro-/anti-inflammatory (M1/M2) ratio 18. Furthermore, it is also able to scavenge the superoxide ion (O2−•) 19 and other reactive species 20.
Amyloid-beta (Aβ) is a 42 amino acids long (amyloid-beta (Aβ)1-42) peptide physiologically present in the brain and cerebrospinal fluid of human beings 21. Together with marked inflammation 22, both the extracellular deposition of insoluble aggregates of this peptide in the brain and its blood vessels 23 and the formation of neurofibrillary tangles composed by the highly phosphorylated form of tau protein 24 represent the neuropathological hallmarks of Alzheimer’s disease (AD). Amyloid-beta (Aβ) peptide can undergo aggregation through a step-by-step process, starting with soluble monomers and evolving to the formation of oligomers, protofibrils, and mature fibrils 25, with the oligomeric structures representing the more toxic species 26.
Microglia are a subtype of brain glial cells and constitute up to 10% of all cells in the healthy human cortex 27. These cells are in intimate contact with neurons and are involved in many basic physiological processes 28. Microglial cells, often found near amyloid-beta (Aβ) plaques in Alzheimer’s disease patients 29, are able to produce various neurotrophic and anti-inflammatory factors essential for cell growth and protection; they can also release different cytotoxic substances, such as reactive oxygen species (ROS) and reactive nitrogen species, and pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α 28. All the above substances are strongly connected to Alzheimer’s disease pathogenesis and amyloid-related neurodegeneration 30.
Nitric oxide (reactive nitrogen species) and O2−• (reactive oxygen species) are part of the natural aerobic metabolism of cells and are involved in many physiological and pathological processes 31. Peroxynitrite, the reaction product between nitric oxide (NO) and O2−•, can react and consequently damage fatty acids, proteins, DNA, and mitochondria leading to oxidative and nitrosative stress 32, inflammation 33, and then neurodegenerative phenomena 34. During the inflammation process, both inducible nitric oxide synthase (iNOS), responsible for nitric oxide (NO) production 35, and NADPH oxidase (Nox), responsible for O2−• production 36, are overactivated in immune cells such as macrophage and microglia 37. When simultaneously activated, inducible nitric oxide synthase (iNOS) and Nox act synergistically to promote neuronal death through the generation of peroxynitrite, which is a more dangerous species if compared with NO and/or O2−• 38. Mediators of cytotoxicity released from activated microglia also include arachidonic acid, glutamate, and histamine 39. Furthermore, it has been shown that macrophages and microglia play a crucial role in several diseases characterized by oxidative stress and inflammation and also that the modulation of their pro-/anti-inflammatory (M1/M2) ratio and secretion products might represent a novel pharmacological approach for the treatment of these disorders 40.
Neuroinflammation is a widely accepted factor associated with the pathogenesis of Alzheimer’s disease 41. Different cell types, including microglia, shift from the resting to activated state during the neuroinflammation process 42, producing a higher amount of pro-inflammatory cytokines such as TNF-α, IL-1β, and IL-6 30. Activated microglia have been shown to contribute to the development and progression of neurodegenerative disorders 43 and the presence of both reactive microglia and astrocytes has been observed in association with amyloid accumulation in Alzheimer’s disease brain 44. However, activated microglia are also able to produce anti-inflammatory mediators 45. Among these molecules, the multifunctional cytokine TGF-β1 has been shown to play a pivotal role in Alzheimer’s disease, exerting neuroprotective effects against amyloid-beta (Aβ)-induced neurodegeneration 46. Furthermore, the secretion of TGF-β1 from peripheral blood mononuclear cells in the circulation 47 along with the levels of TGF-β1 in the plasma 48 are reduced in Alzheimer’s disease subjects. Lastly, Wyss-Coray et al. 49 showed that TGF-β1 promoted microglial amyloid-beta (Aβ) clearance and the reduction of plaque burden in Alzheimer’s disease mice and enhanced amyloid-beta (Aβ) clearance by BV-2 microglial cells, suggesting a link between microglia activity, TGF-β1 release, and the neuroprotective activity of this neurotrophic factor against amyloid-beta (Aβ)-induced toxicity.
In this study 2, Caruso and colleagues reported for the first time that carnosine prevents Aβ-induced oxidative stress in BV-2 microglial cells by decreasing the expression of inducible nitric oxide synthase and NADPH oxidase and the concentrations of nitric oxide and superoxide anion. The authors demonstrated for the first time that, in an established model of Aβ-induced inflammation, carnosine was able to decrease the secretion of pro-inflammatory cytokines such as IL-1β, simultaneously rescuing IL-10 levels and increasing the synthesis and the release of TGF-β1. The authors then validated the protective activity of carnosine in mixed neuronal cultures challenged with Aβ1-42 oligomers, where carnosine prevented Aβ-induced neurodegenerative phenomena via the activation of TGF-β1 signaling.
The inhibition of Aβ oligomer-mediated inflammation and rescue of TGF-β1 signaling have been recently considered effective strategies for protecting against neurodegeneration and disease progression in Alzheimer’s disease 2. Carnosine, through its multimodal mechanism of action, might represent a new pharmacological tool to yield neuroprotection in Alzheimer’s disease 2.
In a double-blind placebo-controlled study addressing the therapeutic potential of carnosine in Alzheimer’s disease, it was assessed the efficacy of a combination of carnosine plus other antioxidants (formula F) in volunteers receiving donepezil (n = 26), in comparison to patients receiving donepezil plus placebo (n = 26) 50. Using the Mini-Mental State Examination II (MMSE II) score, Cornelli 50 observed a significantly improvement of cognition in the group receiving donepezil plus formula F, as compared to those receiving donepezil plus placebo.
A pilot study performed by Boldyrev and colleagues 51 assessed the efficacy of carnosine (1.5 g, daily, by 30 days) as a complement to DOPA (levodopa) drug therapy in Parkinson’s disease patients. It was concluded that the combination of carnosine with basic therapy for certain Parkinson’s disease patients may be a reasonable way to improve the protocol of Parkinson’s disease treatment and to decrease the possible toxic effects of the overloading of DOPA-containing drugs 51. Due to the small number of volunteers and the short duration of this study, a larger and long-term trial is needed to confirm the benefit of carnosine combined to levodopa drug therapy.
L-carnosine and autism
Little is known about the effects of carnosine supplementation for seizures in autism spectrum disorder 52. Some autistic patients take carnosine as a dietary supplement and attribute an improvement in their condition to it. L-carnosine has been demonstrated to significantly improve the Gilliam Autism Rating Scale and the Receptive One-Word Picture Vocabulary tests without any adverse effects in an 8-week double-blind, placebo-controlled study in children with autism spectrum disorder 53. With only one l-carnosine prospective, non-randomized study for the treatment of cognitive and behavioral symptoms of autism spectrum disorder.
Supplemental carnosine may increase corticosterone levels. This may explain the “hyperactivity” seen in autistic subjects at higher doses.
Carnosine eye drops
Cataract is associated with conformational changes and unfolding of proteins in the lens, which could arise from post-translational modifications induced by the Maillard reaction 54. This process is defined as a non-enzymatic modification of amino acids by reaction with sugars, cyanide or steroids. The unfolded proteins of the lens are susceptible to oxidative damage, characterized by the linkage of polypeptide chains through disulphide bonds and formation of high molecular weight water-insoluble aggregates. Such molecules account for the increased scattering of light, resulting in the opacity of the lens typical of the cataract 55.
In this scenario, an in vitro study showed that the dipeptides N-acetylcarnosine – a pro-drug version of carnosine (N-acetyl-β-alanyl-L-histidine) – and anserine, but not carnosine itself, significantly suppressed UV-induced aggregation of the β-L-crystallin – a protein associated to the lens. An inhibition at the initial stages of protein photochemical damage and a decrease in size of protein aggregates were also detected in the presence of both dipeptides 56. In vivo and ex vivo studies also showed the antioxidant activity of the carnosine, N-acetylcarnosine, and β-alanyl-L-histamine. These compounds significantly prevented biological molecules from lipid peroxide-induced damage, including proteins, enzymes, unsaturated fatty acids, and membrane phospholipids, which may occur in the lens cells and predispose to cataract 57. Importantly, the proposed mechanism of action for N-acetylcarnosine is the anti-lipid peroxidation 55, representing a modified form of carnosine resistant to hydrolysis by the human serum carnosinase 58.
Ophthalmic formulations containing N-acetylcarnosine may be promising for treating many ocular diseases mediated by lipid peroxide- and ROS-induced damage, including cataract 59, since such disorder is associated with the lack of a reductive detoxification system of phospholipid hydroperoxides in the lens cell membranes and biomolecules 60. Telomere shortening and increased oxidative stress in human lens cells may be the result of the peroxidative damage to such cells, which is associated with the cause of cataract. Therefore, telomere shortening rate as well as damages to telomere of the lens cells could be reduced by the systemic administration of carnosine (protected from enzyme hydrolysis), N-acetylcarnosine eye drops, and ophthalmic formulations 61.
By convention, a suitable clinical trial of any drug should be performed with a large number of volunteers and by a long-term as a multicentre, randomized, double blind, and placebo-controlled trial. In spite of the promising evidences, the clinical studies performed to date with N-acetylcarnosine eye drops 62 present certain limitations, including a limited number of patients. Further independent research of clinical effectiveness is needed to validate research claims of N-acetylcarnosine that have potential conflicts of interest in humans 63.
Carnosine foods
Foods such as meats or fish contain considerable amounts of carnosine 64.
Carnosine supplement
Carnosine supplements are generally available in tablets at strengths between 100 and 500 mg. There are questions about the efficacy of oral supplementation with carnosine. Most carnosine absorbed from the gut is destroyed in the bloodstream by enzymes called carnosinases. These enzymes readily split carnosine back into its two constituent amino acids, beta-alanine and histidine. Unmetabolized portions enter systemic circulation and are gradually transported into the muscles and other tissues. At the same time, carnosine is also hydrolyzed in the kidneys and excreted. Thus, although specific half-life of carnitine has not been established, it is presumed to be short, and intravenous doses have shown excretion after 90 minutes. This fact has led to trials of intranasal administration of carnosine. This might be a very clever way to get it into areas of the brain, such as the olfactory lobes, that are ravaged by glycosylation and oxidative damage in sufferers of Alzheimer’s Disease. Nonetheless, there is evidence from human studies that supplementing with carnosine can increase carnosine levels in muscle tissue. Supplying people with the amino acids the body uses to make carnosine also increases carnosine levels in muscle. Therefore, even if enzymes do break carnosine apart in the blood, the pieces can be put back together in the cells.
Futhermore, there is little evidence from human studies examining the efficacy of carnosine supplementation in reducing cardiometabolic risk factors 65.
Lastly, unless you are practicing a strict vegetarian diet, carnosine supplementation is likely unnecessary. Carnosine levels in the blood increase after a person has a meal of beef. Whereas a 1000 mg of carnosine a day has been recommended as a supplement, there is about 1500 mg of carnosine in a pound of beef, and close to 2000 mg in similar amounts of pork or chicken. Most fish, such as salmon, are low in carnosine, but high in a substance called anserine. Anserine is also found in the human body, and has actions in cells quite similar to those of carnosine 3.
Carnosine supplement side effects
Generally carnosine is well tolerated and no adverse reactions have been reported. Similarly, no upper limits of intake have been established, and overdosage has been reported.
- Hipkiss, A.R.; Brownson, C.; Bertani, M.F.; Ruiz, E.; Ferro, A. Reaction of carnosine with aged proteins: Another protective process? Ann. N. Y. Acad. Sci. 2002, 959, 285–294.[↩]
- Carnosine Prevents Aβ-Induced Oxidative Stress and Inflammation in Microglial Cells: A Key Role of TGF-β1. Cells 2019, 8(1), 64; doi:10.3390/cells8010064 https://www.mdpi.com/2073-4409/8/1/64/htm[↩][↩][↩][↩]
- NUTRITIONAL SUPPLEMENTS AND METABOLIC SYNDROME. Metabolic Syndrome and Psychiatric Illness. Interactions, Pathophysiology, Assessment and Treatment 2008, Pages 141-186 https://doi.org/10.1016/B978-012374240-7.50012-7[↩][↩][↩]
- HIPKISS, A. R. (2006), Would Carnosine or a Carnivorous Diet Help Suppress Aging and Associated Pathologies?. Annals of the New York Academy of Sciences, 1067: 369-374. doi:10.1196/annals.1354.052 https://doi.org/10.1196/annals.1354.052[↩][↩]
- Hipkiss, A.R. On the relationship between energy metabolism, proteostasis, aging and Parkinson’s disease: Possible causative role of methylglyoxal and alleviative potential of carnosine. Aging Dis. 2017, 8, 334–345.[↩]
- Serum carnosinase deficiency concomitant with mental retardation. Murphey WH, Lindmark DG, Patchen LI, Housler ME, Harrod EK, Mosovich L. Pediatr Res. 1973 Jul; 7(7):601-6.[↩][↩][↩]
- Pearl PL,Jakobs C,Gibson KM (2007). Disorders of β- and γ-amino acids in free and peptide-linked forms. In:Scriver CR,Sly WS,Childs B, editors. The Online Metabolic & Molecular Bases of Inherited Disease. New York: McGraw-Hill.[↩]
- A deletion in the long arm of chromosome 18 in a child with serum carnosinase deficiency. Willi SM, Zhang Y, Hill JB, Phelan MC, Michaelis RC, Holden KR. Pediatr Res. 1997 Feb; 41(2):210-3.[↩]
- Homology modeling of human serum carnosinase, a potential medicinal target, and MD simulations of its allosteric activation by citrate. Vistoli G, Pedretti A, Cattaneo M, Aldini G, Testa B. J Med Chem. 2006 Jun 1; 49(11):3269-77.[↩]
- Janssen B,Hohenadel D,Brinkkoetter P,Peters V,Rind N,Fischer C, et al. (2005). Carnosine as a protective factor in diabetic nephropathy: association with a leucine repeat of the carnosinase gene CNDP1. Diabetes, 54:2320-7[↩]
- Gjessing LR,Lunde HA,Mørkrid L,Lenney JF,Sjaastad Ø (1990). Inborn errors of carnosine and homocarnosine metabolism. J Neural Transm Suppl, 29:91-106.[↩][↩]
- Lenney JF,Peppers SC,Kucera CM,Sjaastad Ø (1983). Homocarnosinosis: lack of serum carnosinase is the defect probably responsible for elevated brain and CSF homocarnosine. Clin Chim Acta, 132:157-65.[↩]
- Physiology and Pathophysiology of Carnosine. Alexander A. Boldyrev, Giancarlo Aldini, and Wim Derave. Physiological Reviews 2013 93:4, 1803-1845 https://doi.org/10.1152/physrev.00039.2012[↩]
- Carnosine may reduce lung injury caused by radiation therapy. Med Hypotheses. 2006;66(5):957-9. Epub 2006 Jan 10. https://www.ncbi.nlm.nih.gov/pubmed/16406688[↩]
- Kulebyakin, K.; Karpova, L.; Lakonsteva, E.; Krasavin, M.; Boldyrev, A. Carnosine protects neurons against oxidative stress and modulates the time profile of mapk cascade signaling. Amino Acids 2012, 43, 91–96.[↩]
- Corona, C.; Frazzini, V.; Silvestri, E.; Lattanzio, R.; La Sorda, R.; Piantelli, M.; Canzoniero, L.M.; Ciavardelli, D.; Rizzarelli, E.; Sensi, S.L. Effects of dietary supplementation of carnosine on mitochondrial dysfunction, amyloid pathology, and cognitive deficits in 3xtg-ad mice. PLoS ONE 2011, 6, e17971.[↩]
- Aloisi, A.; Barca, A.; Romano, A.; Guerrieri, S.; Storelli, C.; Rinaldi, R.; Verri, T. Anti-aggregating effect of the naturally occurring dipeptide carnosine on abeta1-42 fibril formation. PLoS ONE 2013, 8, e68159[↩]
- Caruso, G.; Fresta, C.G.; Martinez-Becerra, F.; Antonio, L.; Johnson, R.T.; de Campos, R.P.S.; Siegel, J.M.; Wijesinghe, M.B.; Lazzarino, G.; Lunte, S.M. Carnosine modulates nitric oxide in stimulated murine raw 264.7 macrophages. Mol. Cell. Biochem. 2017, 431, 197–210.[↩]
- Guliaeva, N.V. [Superoxide-scavenging activity of carnosine in the presence of copper and zinc ions]. Biokhimiia 1987, 52, 1216–1220[↩]
- Cripps, M.J.; Hanna, K.; Lavilla, C., Jr.; Sayers, S.R.; Caton, P.W.; Sims, C.; De Girolamo, L.; Sale, C.; Turner, M.D. Carnosine scavenging of glucolipotoxic free radicals enhances insulin secretion and glucose uptake. Sci. Rep. 2017, 7, 13313.[↩]
- Haass, C.; Schlossmacher, M.G.; Hung, A.Y.; Vigo-Pelfrey, C.; Mellon, A.; Ostaszewski, B.L.; Lieberburg, I.; Koo, E.H.; Schenk, D.; Teplow, D.B.; et al. Amyloid beta-peptide is produced by cultured cells during normal metabolism. Nature 1992, 359, 322–325.[↩]
- Sastre, M.; Klockgether, T.; Heneka, M.T. Contribution of inflammatory processes to alzheimer’s disease: Molecular mechanisms. Int. J. Dev. Neurosci. 2006, 24, 167–176.[↩]
- ounkin, S.G. Evidence that a beta 42 is the real culprit in alzheimer’s disease. Ann. Neurol. 1995, 37, 287–288.[↩]
- Brion, J.P. Neurofibrillary tangles and alzheimer’s disease. Eur. Neurol. 1998, 40, 130–140.[↩]
- Brorsson, A.C.; Kumita, J.R.; MacLeod, I.; Bolognesi, B.; Speretta, E.; Luheshi, L.M.; Knowles, T.P.; Dobson, C.M.; Crowther, D.C. Methods and models in neurodegenerative and systemic protein aggregation diseases. Front. Biosci. 2010, 15, 373–396[↩]
- Selkoe, D.J. Soluble oligomers of the amyloid beta-protein impair synaptic plasticity and behavior. Behav. Brain Res. 2008, 192, 106–113[↩]
- Mittelbronn, M.; Dietz, K.; Schluesener, H.J.; Meyermann, R. Local distribution of microglia in the normal adult human central nervous system differs by up to one order of magnitude. Acta Neuropathol. 2001, 101, 249–255.[↩]
- Kraft, A.D.; Harry, G.J. Features of microglia and neuroinflammation relevant to environmental exposure and neurotoxicity. Int. J. Environ. Res. Public Health 2011, 8, 2980–3018.[↩][↩]
- Perlmutter, L.S.; Scott, S.A.; Barron, E.; Chui, H.C. Mhc class ii-positive microglia in human brain: Association with alzheimer lesions. J. Neurosci. Res. 1992, 33, 549–558.[↩]
- Wang, W.Y.; Tan, M.S.; Yu, J.T.; Tan, L. Role of pro-inflammatory cytokines released from microglia in alzheimer’s disease. Ann. Transl. Med. 2015, 3, 136.[↩][↩]
- De Campos, R.P.; Siegel, J.M.; Fresta, C.G.; Caruso, G.; da Silva, J.A.; Lunte, S.M. Indirect detection of superoxide in raw 264.7 macrophage cells using microchip electrophoresis coupled to laser-induced fluorescence. Anal. Bioanal. Chem. 2015, 407, 7003–7012[↩]
- Maes, M.; Galecki, P.; Chang, Y.S.; Berk, M. A review on the oxidative and nitrosative stress (O&NS) pathways in major depression and their possible contribution to the (NEURO)degenerative processes in that illness. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 676–692.[↩]
- Fubini, B.; Hubbard, A. Reactive oxygen species (ROS) and reactive nitrogen species (RNS) generation by silica in inflammation and fibrosis. Free Radic. Biol. Med. 2003, 34, 1507–1516.[↩]
- Kim, G.H.; Kim, J.E.; Rhie, S.J.; Yoon, S. The role of oxidative stress in neurodegenerative diseases. Exp. Neurobiol. 2015, 24, 325–340.[↩]
- Metto, E.C.; Evans, K.; Barney, P.; Culbertson, A.H.; Gunasekara, D.B.; Caruso, G.; Hulvey, M.K.; Fracassi da Silva, J.A.; Lunte, S.M.; Culbertson, C.T. An integrated microfluidic device for monitoring changes in nitric oxide production in single t-lymphocyte (jurkat) cells. Anal. Chem. 2013, 85, 10188–10195[↩]
- Sorescu, D.; Weiss, D.; Lassegue, B.; Clempus, R.E.; Szocs, K.; Sorescu, G.P.; Valppu, L.; Quinn, M.T.; Lambeth, J.D.; Vega, J.D.; et al. Superoxide production and expression of nox family proteins in human atherosclerosis. Circulation 2002, 105, 1429–1435.[↩]
- Siegel, J.M.; Schilly, K.M.; Wijesinghe, M.B.; Caruso, G.; Fresta, C.G.; Lunte, S.M. Optimization of a microchip electrophoresis method with electrochemical detection for the determination of nitrite in macrophage cells as an indicator of nitric oxide production. Anal. Methods 2019, 11, 148–156.[↩]
- Beckman, J.S.; Crow, J.P. Pathological implications of nitric oxide, superoxide and peroxynitrite formation. Biochem. Soc. Trans. 1993, 21, 330–334.[↩]
- Nakajima, K.; Kohsaka, S. Microglia: Activation and their significance in the central nervous system. J. Biochem. 2001, 130, 169–175.[↩]
- Lucherini, O.M.; Lopalco, G.; Cantarini, L.; Emmi, G.; Lopalco, A.; Venerito, V.; Vitale, A.; Iannone, F. Critical regulation of Th17 cell differentiation by serum amyloid-A signalling in Behcet’s disease. Immunol Lett. 2018, 201, 38–44.[↩]
- Heneka, M.T.; O’Banion, M.K.; Terwel, D.; Kummer, M.P. Neuroinflammatory processes in alzheimer’s disease. J. Neural Transm. 2010, 117, 919–947.[↩]
- Rojo, L.E.; Fernandez, J.A.; Maccioni, A.A.; Jimenez, J.M.; Maccioni, R.B. Neuroinflammation: Implications for the pathogenesis and molecular diagnosis of alzheimer’s disease. Arch. Med. Res. 2008, 39, 1–16.[↩]
- Smith, J.A.; Das, A.; Ray, S.K.; Banik, N.L. Role of pro-inflammatory cytokines released from microglia in neurodegenerative diseases. Brain Res. Bull. 2012, 87, 10–20.[↩]
- Schwab, C.; McGeer, P.L. Inflammatory aspects of alzheimer disease and other neurodegenerative disorders. J. Alzheimers Dis. 2008, 13, 359–369[↩]
- Merlo, S.; Spampinato, S.F.; Beneventano, M.; Sortino, M.A. The contribution of microglia to early synaptic compensatory responses that precede β-amyloid-induced neuronal death. Sci. Rep. 2018, 8, 7297.[↩]
- Caraci, F.; Spampinato, S.F.; Morgese, M.G.; Tascedda, F.; Salluzzo, M.G.; Giambirtone, M.C.; Caruso, G.; Munafo, A.; Torrisi, S.A.; Leggio, G.M.; et al. Neurobiological links between depression and ad: The role of tgf-beta1 signaling as a new pharmacological target. Pharmacol. Res. 2018, 130, 374–384.[↩]
- Luppi, C.; Fioravanti, M.; Bertolini, B.; Inguscio, M.; Grugnetti, A.; Guerriero, F.; Rovelli, C.; Cantoni, F.; Guagnano, P.; Marazzi, E.; et al. Growth factors decrease in subjects with mild to moderate alzheimer’s disease (AD): Potential correction with dehydroepiandrosterone-sulphate (DHEAS). Arch. Gerontol. Geriatr. 2009, 49 (Suppl. 1), 173–184.[↩]
- Juraskova, B.; Andrys, C.; Holmerova, I.; Solichova, D.; Hrnciarikova, D.; Vankova, H.; Vasatko, T.; Krejsek, J. Transforming growth factor beta and soluble endoglin in the healthy senior and in alzheimer’s disease patients. J. Nutr. Health Aging 2010, 14, 758–761[↩]
- Wyss-Coray, T.; Lin, C.; Yan, F.; Yu, G.Q.; Rohde, M.; McConlogue, L.; Masliah, E.; Mucke, L. TGF-beta1 promotes microglial amyloid-beta clearance and reduces plaque burden in transgenic mice. Nat. Med. 2001, 7, 612–618.[↩]
- Treatment of Alzheimer’s disease with a cholinesterase inhibitor combined with antioxidants. Cornelli U. Neurodegener Dis. 2010; 7(1-3):193-202. https://www.ncbi.nlm.nih.gov/pubmed/20224285/[↩][↩]
- Carnosine [corrected] increases efficiency of DOPA therapy of Parkinson’s disease: a pilot study. Boldyrev A, Fedorova T, Stepanova M, Dobrotvorskaya I, Kozlova E, Boldanova N, Bagyeva G, Ivanova-Smolenskaya I, Illarioshkin S. Rejuvenation Res. 2008 Aug; 11(4):821-7. https://www.ncbi.nlm.nih.gov/pubmed/18729814/[↩][↩]
- Frye RE, Rossignol D, Casanova MF, et al. A review of traditional and novel treatments for seizures in autism spectrum disorder: findings from a systematic review and expert panel. Front Public Health. 2013;1:31. Published 2013 Sep 13. doi:10.3389/fpubh.2013.00031 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3859980/[↩]
- Double-blind, placebo-controlled study of L-carnosine supplementation in children with autistic spectrum disorders. Chez MG, Buchanan CP, Aimonovitch MC, Becker M, Schaefer K, Black C, Komen J. J Child Neurol. 2002 Nov; 17(11):833-7.[↩]
- Crabbe MJ (1998). Cataract as a conformational disease – the Maillard reaction, α-crystallin and chemotherapy. Cell Mol Biol (Noisy-le-grand), 44:1047-50[↩]
- Toh T,Morton J,Coxon J,Elder MJ (2007). Medical treatment of cataract. Clin Experiment Ophthalmol, 35:664-71.[↩][↩]
- Dizhevskaya AK,Muranov KO,Boldyrev AA,Ostrovsky MA (2012). Natural dipeptides as mini-chaperones: molecular mechanism of inhibition of lens βL-crystallin aggregation. Curr Aging Sci, 5:236-41.[↩]
- Babizhayev MA (2006). Biological activities of the natural imidazole-containing peptidomimetics N-acetylcarnosine, carcinine and L-carnosine in ophthalmic and skin care products. Life Sci, 78:2343-57.[↩]
- Hipkiss AR,Cartwright SP,Bromley C,Gross SR,Bill RM (2013). Carnosine: can understanding its actions on energy metabolism and protein homeostasis inform its therapeutic potential? Chem Cent J, 7:38[↩]
- Babizhayev MA (2011). Mitochondria induce oxidative stress, generation of reactive oxygen species and redox state unbalance of the eye lens leading to human cataract formation: disruption of redox lens organization by phospholipid hydroperoxides as a common basis for cataract disease. Cell Biochem Funct, 29:183-206.[↩]
- Babizhayev MA (1996). Failure to withstand oxidative stress induced by phospholipid hydroperoxides as a possible cause of the lens opacities in systemic diseases and ageing. Biochim Biophys Acta, 1315:87-99[↩]
- Babizhayev MA,Vishnyakova KS,Yegorov YE (2011). Telomere-dependent senescent phenotype of lens epithelial cells as a biological marker of aging and cataractogenesis: the role of oxidative stress intensity and specific mechanism of phospholipid hydroperoxide toxicity in lens and aqueous. Fundam Clin Pharmacol, 25:139-62[↩]
- Babizhayev MA,Micans P,Guiotto A,Kasus-Jacobi A (2009). N-Acetylcarnosine lubricant eyedrops possess all-in-one universal antioxidant protective effects of L-carnosine in aqueous and lipid membrane environments, aldehyde scavenging, and transglycation activities inherent to cataracts: a clinical study of the new vision-saving drug N-acetylcarnosine eyedrop therapy in a database population of over 50,500 patients. Am J Ther, 16:517-33.[↩]
- Cloud A,Tandon A,Calhoun J,Cebulla CM (2012). Rapid formation and resolution of cataracts following orthopedic surgery for a patient with Charcot-Marie-Tooth disease. Arch Ophthalmol, 130:260-2.[↩]
- Zinc, Carnosine, and Neurodegenerative Diseases. Nutrients 2018, 10(2), 147; doi:10.3390/nu10020147 https://www.mdpi.com/2072-6643/10/2/147/htm[↩]
- Elbarbary, N.S.; Ismail, E.A.R.; El-Naggar, A.R.; Hamouda, M.H.; El-Hamamsy, M. The effect of 12 weeks carnosine supplementation on renal functional integrity and oxidative stress in pediatric patients with diabetic nephropathy: A randomized placebo-controlled trial. Pediatr. Diabetes 2017, 19, 470–477.[↩]

{kind=link}