What is lactoferrin
Lactoferrin is a multifunctional cationic glycoprotein of 691 amino acids constitutively synthesized by exocrine glands and by neutrophils following infection and inflammation, is present in human fluids 1. Lactoferrin was first identified in 1939 in bovine milk and isolated in 1960 from both human 2 and bovine milk 3. Human colostrum shows the highest levels of lactoferrin (~7 g/L) 4, while mature milk, other secretions and secondary granules of neutrophils present lower levels (Table 1) 5. During infection and/or inflammation processes, the lactoferrin concentration increases through the recruitment of neutrophils. Remarkably, 106 neutrophils synthesize 15 μg of lactoferrin 1.
Table 1. Lactoferrin concentration in different human fluids and secretions
| Biological Fluids | Concentration (mg/mL) |
|---|---|
| Colostrum | 8 |
| Milk | 1.5–4 |
| Tears | 2 |
| Saliva | 0.008 |
| Vaginal secretion | 0.008 |
| Seminal fluid | 0.112 |
| Cerebrospinal fluid | Undetectable |
| Plasma | 0.0004 |
| Joint fluid | 0.001 |
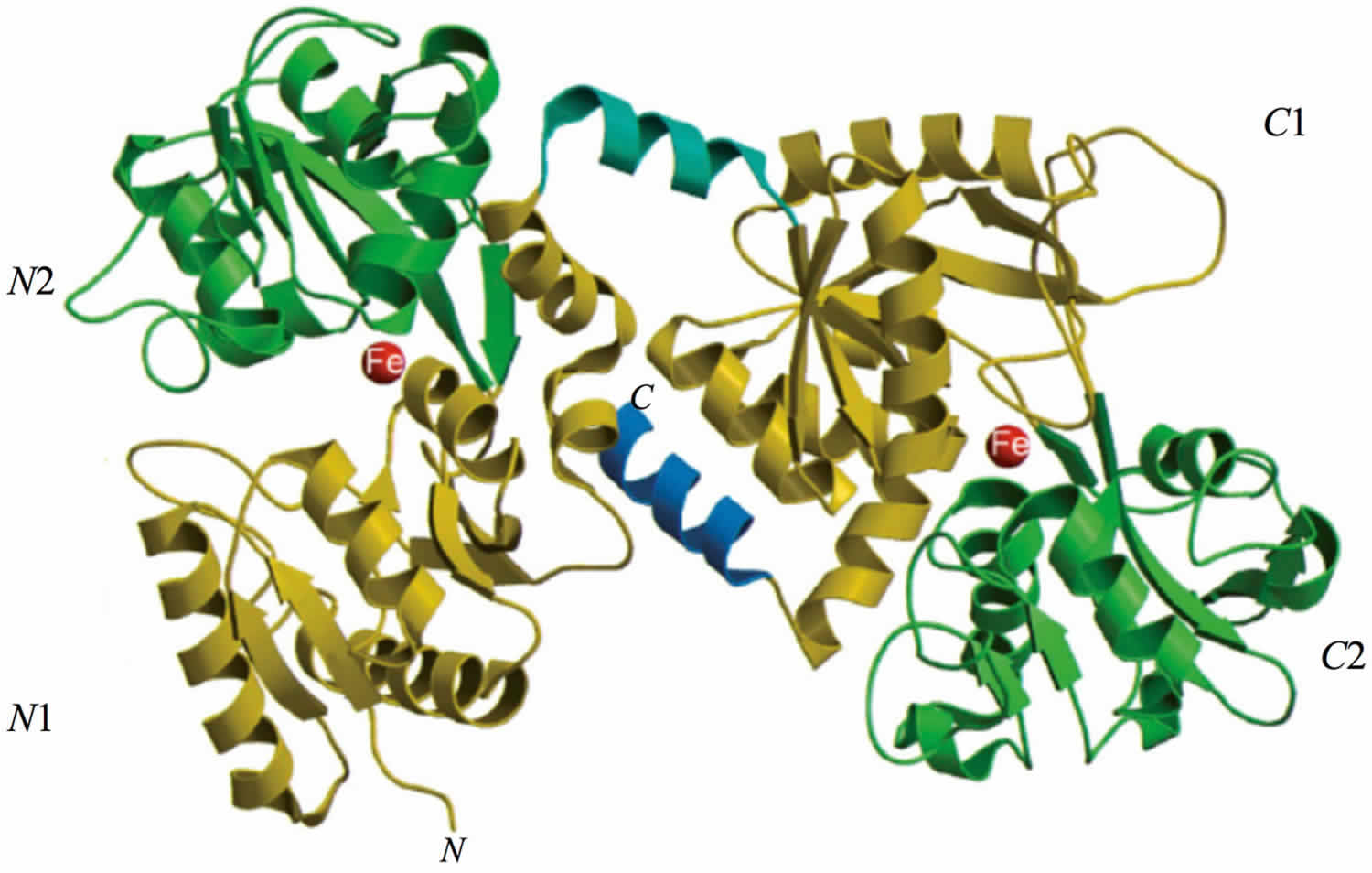
Human lactoferrin is divided into two homologous lobes (N-lobe residues 1–333 and C-lobe residues 345–691) connected by a 3-turn α-helix peptide (residues 334–344) 6. Each lobe, constituted by two domains (N1 and N2, C1 and C2), binds one ferric ion and one carbonate anion within a deep cleft between the domains of each lobe (Figure 1) 6. All lactoferrins can adopt two main conformational states: the open metal-free (apo-lactoferrin) and the closed metal-bound (holo-lactoferrin). Metal binding and release are thus associated with large-scale conformational changes in which the domains close over the bound metal ion or open to release it 7. The iron-saturated closed form is highly stable and more resistant to digestion by proteases compared to the unsaturated open one 8. Within the iron-binding transferrin family, lactoferrin is able to reversibly chelate two Fe3+ ions per molecule with high affinity, as well as retain ferric iron until pH values as low as 3.0, characteristic of the infection and inflammation sites. Of note, the iron binding ability of lactoferrin in secretions and of transferrin in cells and circulation guarantees that free available iron does not exceed 10−18 M, thus preventing (i) iron precipitation as insoluble hydroxides, (ii) microbial growth and (iii) the formation of ROS (reactive oxygen species), responsible for tissue, cell, DNA, protein and membrane lipid damage. In addition to the iron-binding ability of lactoferrin and transferrin, the closed forms can include other transition metal ions such as copper ion (Cu2+) and manganese ion (Mn3+), chelated at lower affinity than Fe3+ without changing the basic structure 9. Conversely, the differences in iron-releasing ability are related to their different functions: anti-microbial, anti-inflammatory and immunomodulatory activities for lactoferrin and iron delivery activity for transferrin.
Figure 1. Structure of lactoferrin
Footnote: The N-lobe on the left and the C-lobe on the right are divided into four domains, labeled N1, N2, C1, C2. The red spheres represent the two ferric ions in each iron-binding site.
[Source 1 ]
Lactoferrin is glycoprotein released by a type of white blood cell called a neutrophil. When there is inflammation in the digestive tract, neutrophils are attracted to the area and release lactoferrin, increasing the level of the glycoprotein in the stool. A positive stool lactoferrin is indicative of the presence of intestinal inflammation (a marker for fecal white blood cells or leukocytes). But a negative result does not exclude the presence of intestinal inflammation.
Intestinal inflammation is associated with inflammatory bowel disease (IBD) and with some bacterial infections of the digestive tract, but it is not associated with many other disorders that affect bowel function and cause similar symptoms. Lactoferrin can be used to help distinguish between inflammatory and non-inflammatory conditions.
Inflammatory bowel disease (IBD) is a group of chronic disorders characterized by swollen and damaged tissues in the lining of the intestinal tract. The cause of inflammatory bowel disease (IBD) is not known, but these diseases are thought to be due to an autoimmune process that has been triggered by a genetic predisposition, a viral illness, and/or an environmental factor. The most common inflammatory bowel diseases are Crohn disease and ulcerative colitis.
People affected by inflammatory bowel disease (IBD) typically have flare-ups of active disease that alternate with periods of remission. During a flare-up, a person may experience frequent bouts of watery and/or bloody diarrhea, abdominal pain, weight loss, and fever. Between these flare-ups, symptoms frequently subside. Many people may go through extended periods of remission between flare-ups. Lactoferrin testing can be useful in monitoring inflammatory bowel disease activity.
What is lactoferrin in stool
Lactoferrin in stool is measured as a way to detect inflammation in the intestines. Lactoferrin is a stool test that is used to detect inflammation in the intestines. Intestinal inflammation is associated with, for example, some bacterial infections and, in people with inflammatory bowel disease (IBD), it is associated with disease activity and severity. Lactoferrin in stool test may be ordered to distinguish between inflammatory bowel disease (IBD) and non-inflammatory disorders and to monitor inflammatory bowel disease disease activity. A baby that is being breast-fed could potentially have a false-positive result because of lactoferrin present in the mother’s breast milk.
Lactoferrin is related to another stool test, calprotectin. Both are substances that are released by white blood cells in the stool and are associated with intestinal inflammation. Of the two tests, calprotectin has been the most extensively studied and it is ordered more frequently than lactoferrin. Usually one or the other will be ordered but not both. Current evidence suggests that there is not a significant benefit to performing both tests as they both detect intestinal inflammation. However, calprotectin has been more extensively studied and is ordered more frequently, but in general they are both thought to be useful tests.
A healthcare practitioner may order a lactoferrin test to help investigate the cause of a person’s persistent watery or bloody diarrhea. The test may be ordered along with other stool tests, such as a stool culture to detect a bacterial infection, a test for ova and parasites (O&P), a stool white blood cell test, and/or a fecal occult blood test.
If a healthcare practitioner suspects inflammation, then blood tests that detect inflammation in the body, such as an ESR (erythrocyte sedimentation rate) and/or CRP (C-reactive protein), may also be ordered. Testing is performed both to help determine what is causing a person’s symptoms and to rule out conditions with similar symptoms. This means that additional blood and stool testing may be performed depending on the suspected causes.
If a person has been diagnosed with inflammatory bowel disease, then a lactoferrin test may be ordered to monitor disease activity and to help evaluate its severity. For example, if a person has a moderately elevated lactoferrin, then testing may be repeated several weeks later to see if it has stayed moderately elevated, increased, or returned to normal.
A lactoferrin test may be ordered to help determine whether an endoscopy is indicated if inflammatory bowel disease is suspected. A diagnosis of inflammatory bowel disease is usually confirmed by performing an endoscopy (colonoscopy or sigmoidoscopy) to examine the intestines and by obtaining a small tissue sample (biopsy) to evaluate for inflammation and changes in tissue structures. This testing is invasive and is less likely to be necessary if inflammation is not present.
When is lactoferrin stool test ordered?
A lactoferrin stool test may be ordered when a person has symptoms that suggest that intestinal inflammation may be present and when a healthcare practitioner wants to distinguish between inflammatory bowel disease (IBD) and a non-inflammatory bowel condition.
Signs and symptoms of intestinal inflammation will vary from person to person and over time. They may include one or more of the following:
- Bloody or watery diarrhea
- Abdominal cramps or pain
- Fever
- Weight loss
- Rectal bleeding
- Weakness
Testing for lactoferrin may be performed when a healthcare practitioner wants to determine whether an endoscopy (colonoscopy or sigmoidoscopy) is likely or less likely to be necessary.
When a person has been diagnosed with inflammatory bowel disease (IBD), a lactoferrin test may be ordered whenever a flare-up is suspected, both to confirm disease activity and to evaluate its severity.
What does fecal lactoferrin result mean?
An elevated lactoferrin level in the stool indicates that inflammation is likely present and active in the digestive tract but does not indicate either its location or cause. In general, the degree of elevation is associated with the severity of the inflammation. An endoscopy (colonoscopy or sigmoidoscopy) may be indicated as a follow-up test.
Increases in lactoferrin are seen with inflammatory bowel disease (IBD) but also with other inflammatory conditions, with intestinal bacterial infections, some parasitic infections, and with colon cancer.
In newly diagnosed people with inflammatory bowel disease (IBD), concentrations of lactoferrin may be very high.
A low level of lactoferrin means that a bowel disorder is likely non-inflammatory. Examples of these include diarrhea due to viral digestive tract infections and irritable bowel syndrome (IBS). Unlike inflammatory bowel disease, irritable bowel syndrome (IBS) does not cause inflammation. Rather, it causes cramp-like stomach pains and spasms with bouts of diarrhea and/or constipation. People with low lactoferrin results are less likely to require an endoscopy.
A moderate lactoferrin level may be seen in those with less active inflammatory bowel disease (IBD). The result indicates that there is likely some inflammation present. If a repeat test shows that the lactoferrin has increased, then the person’s condition may be worsening.
What can I do to decrease my lactoferrin?
Lactoferrin is a reflection of intestinal inflammation and is not affected by lifestyle changes. If it is due to an infection, then it will most likely return to normal when the infection resolves. If it is due to inflammatory bowel disease, then it will rise and fall with disease activity.
What is colostrum lactoferrin?
Lactoferrin is a major antimicrobial protein found naturally in human breast milk 10. The antimicrobial activity of lactoferrin comes from its ability to directly kill microbes by depriving microbes of iron by sequestering iron and make it unavailable for microbial growth, or disrupting their plasma membranes through its highly cationic charge 11. Lactoferrin also has anti-inflammatory and immunomodulatory effects within the gut 12. Due to these properties, lactoferrin may significantly decrease the burden of both late onset sepsis and necrotising enterocolitis, when supplemented to preterm neonates by controlling the overgrowth of sepsis causing pathogens and decreasing inflammation in the gut 13.
Preterm neonates are highly susceptible to infectious and inflammatory diseases such as late-onset sepsis and necrotising enterocolitis and suffer high morbidity 14. Gut dysbiosis, a loss of microbial diversity and overgrowth of Proteobacteria in the small intestine 15, can lead to a cascade of inflammation and translocation of pathogens into the bloodstream 16. Dysbiosis of the preterm infant gastrointestinal tract is a major factor contributing to the development of necrotising enterocolitis and late-onset sepsis 17.
Human breast milk feeding is beneficial in preterm neonates and promotes development of a healthy intestinal microbiome, along with passive immune protection via a variety of soluble and cellular components 18. A higher consumption volume of breast milk in preterm neonates correlates with a lowered risk of late onset sepsis and necrotising enterocolitis 19. Ongoing clinical trials, such as the Lactoferrin Infant Feeding Trial and Enteral Lactoferrin in Neonates 13, are evaluating the benefits of high dose supplementation of breast milk with bovine lactoferrin (cow’s milk lactoferrin) to prevent and limit the impact of late onset sepsis and necrotising enterocolitis. Probiotics are beneficial live microbes which when administered can promote microbiota diversity, improve gut barrier function and colonization with other healthy commensals 20. When administered to preterm neonates, probiotics can seed the lower gut and decrease the burden of late onset sepsis and necrotising enterocolitis by decreasing the risk of inflammation, dysbiosis and translocation of pathogens 21.
Colonization with probiotic bacteria, and simultaneous control of the overgrowth of sepsis pathogens in the preterm gut, may be critical for preventing late onset sepsis and for dampening inflammation. Supplementation of enteral feeds with probiotics or bovine lactoferrin has shown positive clinical outcomes for late onset sepsis and necrotising enterocolitis prevention. One clinical trial has combined these two interventions and shown a 4.6% decrease in late onset sepsis cases. There were also no cases of necrotising enterocolitis in this study in the combination treatment group, compared to the 6% incidence in the placebo group 22. Despite these positive clinical outcomes, this trial did not examine whether there was an additive benefit by co-administration of these two supplements.
Lactoferrin function
Human lactoferrin and bovine lactoferrin possess high sequence homology and exert identical multifunctions: antibacterial, antifungal, antiviral and antiparasitic, anti-inflammatory and immunomodulatory activities 23. Therefore, the majority of the in vitro and in vivo studies have been carried out using bovine lactoferrin, generally recognized as a safe substance (GRAS) by the Food and Drug Administration (FDA, USA) and available in large quantities 1. All of the functions ascribed to lactoferrin can be dependent or independent of lactoferrin-iron-binding ability.
Antibacterial and Anti-Biofilm Activity Dependent on lactoferrin Iron-Binding Ability
The first function attributed to human lactoferrin/bovine lactoferrin (lactoferrin) was the antimicrobial activity. The bacteriostatic action of lactoferrin is usually iron dependent, as iron supplementation reverts its effect 24. The lactoferrin antibacterial activity is counteracted by three main mechanisms put in place by bacterial pathogens:
- Synthesis of high affinity ferric ion chelators, named siderophores, that compete with iron-binding proteins for iron acquisition and delivery into bacteria through specific receptors 25;
- Iron acquisition through lactoferrin or transferrin binding mediated by their specific surface receptors 26; iron acquisition through hemoglobin, haptoglobin and hemopexin binding mediated by surface hemoprotein receptors 27; iron acquisition through heme binding mediated by the surface hemophore receptor 28;
- Iron acquisition through bacterial reductase able to reduce ferric to ferrous ions, thus eliminating the substrate of the Fenton reaction and assimilating ferrous ions that passively enter inside microbial cells 29. Singh et al. 30 demonstrated another important iron-dependent lactoferrin function: inhibition of Pseudomonas aeruginosa biofilm formation in cystic fibrosis (CF) by the iron-binding activity of lactoferrin. As a matter of fact, cystic fibrosis is associated with alterations in the influx and efflux of chloride and sodium ions, which involves also abnormal high concentrations of iron and ferritin in sputum 31. This increased availability of iron (median value of 6.3 × 10−5 M) induces the generation of ROS (reactive oxygen species), which contributes to lung disorders, as well as to the enhanced growth and colonization of Pseudomonas aeruginosa and Burkholderia cepacia, two motile Gram-negative pathogens that are a major source of the morbidity and mortality of cystic fibrosis patients. For both bacteria, biofilm formation is one of the major virulence factors. Peptides and proteins of natural non-immune defenses, including lactoferrin, play a crucial role in combating such infections. A striking Singh et al. 30 discovery was that apo-lactoferrin, by chelating iron, inhibits Pseudomonas aeruginosa adhesion and biofilm formation through activation of a specialized form of motility, named switching. Like Pseudomonas aeruginosa, also free-living forms of Burkholderia cepacia show a noticeable motility under iron-limiting conditions. On the other hand, iron availability or the addition of iron-saturated bovine lactoferrin inhibits the motility and induces abundant Pseudomonas aeruginosa and Burkholderia cepacia growth and aggregates, evolving into biofilm 32. In cystic fibrosis patients, however, these protective effects of lactoferrin are compromised by the presence of high iron concentrations and, consequently, by high levels of holo-lactoferrin 33]. Even if the human lactoferrin concentration increases in infection and inflammation processes, in sputum of cystic fibrosis patients, free iron concentrations remain higher than in normal subjects 34. The high iron concentration (6.3 × 10−5 M) saturates hlactoferrin (1 × 10−5 M), thus preventing human lactoferrin from inhibiting biofilm formation.
Antibacterial Activity Independent of lactoferrin Iron-Binding Ability
An iron-independent bactericidal action is exerted by lactoferrin direct interaction with the lipopolysaccharide (LPS) of Gram-negative or with the lipoteichoic acid of Gram-positive bacteria 35. The bactericidal activity of lactoferrin is located in the N-terminal region, as its derivative cationic peptide, generated by pepsin digestion, called lactoferricin (lactoferrincin), is several folds more active than the intact protein in interacting with lipopolysaccharide and in killing Gram-negative bacteria 36. It is also important to underline that the presence of high calcium concentrations can counteract the release of lipopolysaccharide from Gram-negative bacteria induced by lactoferrin. In fact, the ability of lactoferrin to bind Ca2+ through the carboxylate groups of the sialic acid residues present on glycan chains provokes the release of significant amounts of lipopolysaccharide from Gram-negative bacteria, without needing a direct interaction with bacteria 37. The bactericidal activity towards Gram-positive bacteria appears to be related to the same cationic residues involved in the bactericidal activity against Gram-negative bacteria 38.
Inhibition of Bacterial Adhesion on Abiotic and Cell Surfaces
Independently from its iron-binding ability, blactoferrin inhibits the bacterial adhesion to host cells through its competitive binding to host cells and/or to microbial surface components 38. Microbial adhesion and subsequent colonization, resulting in biofilm formation on abiotic surfaces, such as catheters, prosthesis and medical devices, represent a serious problem that can lead to illness and death. Efforts to reduce microbial adhesion, using new materials or compounds inhibiting microbial adhesion, have had modest success once applied to the patient. Consequently, it would be very helpful to discover other compounds able to hinder microbial adhesion. In 1989, the ability of lactoferrin, in both apo- and holo-form, to inhibit the adhesion of Streptococcus mutans to hydroxyapatite (HA), mimicking the tooth surface, was an interesting disclosure 39. The further demonstration that lactoferrin inhibits the adhesion of Streptococcus mutans to hydroxyapatite through residues 473–538 of its C-lobe confirmed that this activity is unrelated to lactoferrin iron-binding properties 40. The influence of lactoferrin on bacterial adhesion on contact lenses has been also shown through the much lower number of adherent P. aeruginosa on human lactoferrin-coated lenses compared to that observed on human lactoferrin non-coated ones 41. The different nature of abiotic surfaces, microbial adhesion mechanisms and in vitro experimental conditions indicate that the inhibition of bacterial adhesion by apo- or holo-lactoferrin can explain the different requirement to exert adhesion: ionic binding to biomaterials, as well as specific binding to bacterial structures, or both.
The ability of microbes to adhere, colonize and form biofilm on host cells is also a crucial step in the development and persistence of infections. The first demonstration of the mucosal protective activity of hlactoferrin against injury by adherent Escherichia coli HB101 was included in the data reported by Longhi et al. 42. Later, it was confirmed that lactoferrin can inhibit the first step for bacterial pathogenesis through the inhibition of bacterial adherence to host cells 43. Lactoferrin has also been shown to inhibit the adherence of enterotoxigenic E. coli (ETEC) to human epithelial cells and to intestinal mucosa of germfree mice 43, as well as the adhesion of three adhesive diarrheagenic E. coli strains (DAEC), enteroaggregative E. coli (EAEC) 44 and enteropathogenic E. coli (EPEC) 45. Human lactoferrin and bovine lactoferrin, human lactoferrincin and bovine lactoferrincin are all able to bind to Gram-negative and Gram-positive bacterial surfaces 46, as well as to host cells, by binding to glycosaminoglycans 47 and specifically to heparan sulfate 48.
However, lactoferrin can prevent adhesion through other mechanisms. The importance of the sugar residues on lactoferrin is suggested by the observation that whereas native human lactoferrin inhibits Shigella spp. adhesion 49, recombinant hlactoferrin (rhlactoferrin), with different glycosylation, has no effect on Shigella flexneri adhesion to epithelial cells 50. Another paper suggests that hlactoferrin, rhlactoferrin and blactoferrin inhibit the attachment of Helicobacter felix to gastric epithelial cells, probably by interaction between oligomannoside-type glycans of lactoferrin and bacterial adhesins that recognize these residues 51. Although inhibition of bacterial adhesion seems generally to be mediated by lactoferrin binding to both bacterial and host cell surfaces, the surprising discovery of hlactoferrin proteolytic activity 52 provided an additional mechanism to explain lactoferrin anti-adhesive activity. Thus, inhibition by hlactoferrin of the adhesion of enteropathogenic E. coli (EPEC) strains 45, which use a type III secretory system to deliver effector proteins into the host cell, was ascribed to hlactoferrin-mediated degradation of the secreted proteins, EspA, B, D 53, as well as hlactoferrin inhibition of H. influenzae and Aggregatibacter actinomycetemcomitans adhesion to the degradation of two colonization factors and of autotransporter proteins, respectively 54.
Although the experimental conditions of the studies reported were different, the pre-incubation of lactoferrin with host cells seems never to inhibit Gram-positive and Gram-negative bacterial adhesion, suggesting that lactoferrin binding to glycosaminoglycans or heparan sulfate is not crucial. Instead, the inhibition of Gram-positive and Gram- negative bacterial adhesion by lactoferrin seems to require lactoferrin binding to bacteria or a putative lactoferrin-mediated degradation of the adhesins or proteins of the secretory systems. During the adhesion process, bacteria are unable to stimulate the epithelial cell inflammatory responses at significant levels 55.
Inhibition of Bacterial Entry into Host Cells
Some mucosal pathogenic bacteria are capable not only of adhering, but also of entering into non-professional phagocytes, such as epithelial cells. Inside host cells, bacteria are in a protective niche in which they can replicate and persist, thus avoiding host defenses. In addition, antibiotic therapies are not always effective at eradicating intracellular pathogens 56. Virulence determinants, such as surface proteins able to bind host cells, play a key role in the entry process inside the host cells. lactoferrin has been shown to inhibit the entry of Gram-negative and Gram-positive facultative intracellular bacteria. The first demonstration of the inhibition of bacterial invasion by blactoferrin involved E. coli HB101(pRI203), a recombinant strain able to enter inside cells owing to the inv gene located in pRI203 plasmid 42. Bacterial entry into host cells is mediated by the binding of bacterial invasin, a product of the inv gene, to the host integrin receptor. The effectiveness of apo- and holo-blactoferrin and blactoferrincin towards E. coli strain HB101 (pRI203) invasion is correlated with their ability to bind to both cultured cells and the bacterial outer membrane 42. Likewise, for Y. enterocolitica and Y. pseudotuberculosis, grown in conditions allowing maximal invasin synthesis, a 10-fold inhibition of invasion of cultured cells by blactoferrincin was observed 57. It appears that the binding of lactoferrin and lactoferrincin to integrins through the same domains that are targeted by invasin, and to glycosaminoglycans and/or heparan sulfate, can induce a dramatic subversion in bacterial-host cell interaction, thus inhibiting bacterial internalization 58. Similar mechanisms apply to the inhibition of the invasion of the Gram-positive bacteria L. monocytogenes, Streptococcus pyogenes (GAS) and Staphylococcus aureus, i.e., apo- or holo-blactoferrin binding to both bacterial adhesins and host cells 59. The ability of blactoferrin to decrease Streptococcus pyogenes invasion was also confirmed by an in vivo trial carried out on 12 children suffering from pharyngitis and already scheduled for tonsillectomy 59. Although all studies, reported above, have been carried out with different facultative intracellular microorganisms in different in vitro models, lactoferrin, in apo- or holo-form, always exerts an inhibiting activity against the microbial internalization 60. In contrast to inhibition of bacterial adhesion, lactoferrin binding to glycosaminoglycans and/or heparan sulfate of host cells seems crucial in inhibiting bacterial internalization. In the invasion process, the facultative intracellular bacteria induce the up-expression of pro-inflammatory cytokines by infected epithelial cells 61. Blactoferrin can efficiently inhibit the invasion of an obligate intracellular bacteria, as C. trachomatis, as well as counteract the inflammatory process promoted by its intracellular localization 62.
Chlamydia trachomatis, responsible for the most common sexually-transmitted bacterial disease worldwide, is asymptomatic in about 80% of women and causes acute and chronic infections. Unlike acute infections, which can be cured with antibiotics, chronic infections are difficult to eradicate and need prolonged therapies, thus increasing the risk of developing antibiotic resistance 63. Even if novel alternative therapies are needed, the difficulty in finding new agents against Chlamydia trachomatis resides in the complex biphasic developmental life-cycle of this peculiar pathogen: extracellular infectious bodies (elementary bodies, EBs) metabolically inactive, and the intracellular non-infectious bodies (reticulate bodies, RBs), metabolically active. In addition, Chlamydia trachomatis infection induces inflammatory processes. The up-expression of pro-inflammatory cytokines such as TNF-α, IL-1β, IL-6 and IL-8 induces direct damage to genital tissues. A great interest in lactoferrin, considered as a prominent component of the first-line defense of the host against infections and inflammation, has been raised. Recently, the protective effect of blactoferrin against Chlamydia trachomatis infection and inflammation in vitro and in vivo has been demonstrated. A preparation of bovine lactoferrin, iron-saturated at 20%, to consent to further iron chelation, was utilized in in vitro model to check its anti-chlamydial activity 62. The incubation of cell monolayers with blactoferrin before the infection or at the moment of the infection significantly inhibited the adhesion and entry of elementary bodies of Chlamydia trachomatis into epithelial cells. Therefore, the inhibition of C. trachomatis infectivity by blactoferrin was dependent on its interaction with the cell surface and especially with glycosaminoglycans and heparan sulfate proteoglycans 47, which are potential receptors for C. trachomatis adhesion 64. Conversely, the preincubation of blactoferrin with C. trachomatis EBs did not influence its infectivity, supporting the idea that the specific interaction between blactoferrin and epithelial host cells could be the sole pivotal mechanism responsible for the inhibition of C. trachomatis invasion 62. Interestingly, the addition of blactoferrin significantly decreased the IL-8 and IL-6 levels synthetized by C. trachomatis-infected cells. These results, demonstrating once again the ability of blactoferrin to down-regulate pro-inflammatory cytokine synthesis and showing for the first time the protective effects of blactoferrin against C. trachomatis infection, led us to investigate its efficacy also in asymptomatic pregnant women positive for C. trachomatis and with high levels of IL-6 in cervical fluids. In a pilot study, seven out of 176 pregnant women enrolled, showing cervical specimens positive for C. trachomatis, were treated with the intravaginal administration of blactoferrin (100 mg) every 8 h for 30 days. Interestingly, after one month, six out of seven pregnant women were negative for C. trachomatis and showed significant decreased IL-6 levels in their cervical vaginal fluids 62. Similar to what was observed in the in vitro model, intravaginal administration of blactoferrin seems to act by protecting host cells against the adhesion and entry of chlamydial EBs, which are released extracellularly after redifferentiation of RBs to EBs. The simultaneous decrease of IL-6 levels could be a marker for the lack of C. trachomatis EBs infection of host cells due to the presence of blactoferrin. In other words, blactoferrin protects host cells, preventing the early phase of infection by EBs. Therefore, the in vivo anti-chlamydial activity of blactoferrin is related to the protection of host cells against the adhesion to and entry into host cells of C. trachomatis EBs, as well as to its anti-inflammatory function 62.
Inhibition of Viral Infections
The antiviral activity of hlactoferrin was described, for the first time, in mice infected with the polycythemia-inducing strain of the Friend virus complex 65. Since 1994, an effective antiviral activity of both hlactoferrin and blactoferrin during the early stage of infection of enveloped and naked viruses was demonstrated. This activity is mainly due to blactoferrin binding to glycosaminoglycans and heparan sulfate or viral particles or both 66, thus inhibiting viral entry into host cells. Despite the antiviral effect of lactoferrin widely demonstrated in in vitro studies, few clinical trials have been carried out, and the related mechanism of action is still under debate.
Nevertheless, the ability of lactoferrin to exert a potent antiviral activity strengthens the idea that this natural glycoprotein is an important brick in the mucosal wall, effective against viral attacks, and it could be usefully applied as a novel strategy for the treatment of viral infections and of inflammation, the major contributing factor to viral disease severity 67. Epidemiological evidence and clinical observations of infections in humans suggest that different viruses may be associated with different inflammatory responses. Whether or not these differences can be attributed to the viruses themselves or to hosts that are susceptible to severe infection or prone to produce high levels of inflammation with a given virus is still under debate.
Anti-Inflammatory Activity of lactoferrin in Infected and Inflamed Host Cells
As already reported, lactoferrin possesses a potent anti-inflammatory activity able to both modulate the inflammatory response by epithelial cells infected by facultative and obligate intracellular bacteria 62 and revert/attenuate the inflammatory response triggered by Toll-like receptor engagement in antigen-presenting cells 68.
Nevertheless, the literature is full of papers showing contradictory effects of lactoferrin on inflammatory processes in different in vitro cell models. However, a deeper analysis of these conflicting in vitro models revealed how experimental conditions can affect the results. In particular, the experimental cell line models, such as epithelial or phagocytic cells, the different type of infecting agents, such as bacteria or viruses or the stimulation with pathogen-associated molecular patterns (PAMPS), such as flagellin, toxin, peptidoglycan, lipopolysaccharide (LPS), and, finally, the use of lactoferrin from different sources are all important factors that can influence the results 69. It is very important to underline the different inflammatory response by epithelial or macrophagic cells injured by the same stimulus. Epithelial cells are less responsive to bacterial pathogen-associated molecular patterns (PAMPS), such as lipopolysaccharide, compared to the high responsivity of phagocytes 70. In particular, cultured epithelial cells treated with LPS or infected by non-invasive adherent E. coli HB101 synthesize very low levels of pro-inflammatory cytokines, making it difficult to highlight the anti-inflammatory activity of blactoferrin 55. Conversely, the same monolayers infected by invasive E. coli HB101 (pRI203) significantly up-express pro-inflammatory cytokines, the synthesis of which is significantly decreased by blactoferrin 55. These two isogenic E. coli strains express an identical LPS, but E. coli HB101 is only able to adhere to surface cell structures, similarly to commensal bacteria, while E. coli HB101 (pRI203) is capable of entering the host through cells as intracellular pathogenic bacteria. Therefore, the different levels of pro-inflammatory cytokines synthesized by infected epithelial cells are independent of the LPS structure, but strongly dependent on the localization of viable infecting bacteria: adherent or intracellular. It is important to underline that the anti-inflammatory activity of blactoferrin was tested in vitro in different epithelial monolayers untreated or treated with blactoferrin at a concentration <100 µg/mL, which does not inhibit the entry of facultative or obligate intracellular bacterial pathogens. Consequently, the monolayers that were untreated or blactoferrin-treated contain a similar number of intracellular bacteria. If the experiments were not designed as described, the anti-inflammatory activity of blactoferrin would be incorrectly ascribed to the different numbers of intracellular bacteria.
Different epithelial monolayers infected with various facultative or obligate intracellular pathogens were found to up-express pro-inflammatory cytokines. The addition of blactoferrin at 100 µg/mL significantly decreased IL-1β, IL-6, IL-8 and NF-κB levels [30,87,100,106]. Blactoferrin also exerts its anti-inflammatory activity in LPS-inflamed macrophages. Human macrophages, responsive to LPS treatment, up-express IL-6, which is significantly inhibited by blactoferrin, which reduces the pathological inflammation and cell damage, similarly to what was observed in epithelial cells invaded by intracellular bacteria 61.
Of note, the anti-inflammatory activity of lactoferrin had been firstly hypothesized by the demonstration that exogenous blactoferrin is internalized from the apical side of host cells and localized in the nuclei 71. In agreement with the nuclear localization of lactoferrin, in 2008, a very elegant and important paper by Suzuki et al. 72 revealed that the N-lobe or the N1.1 sub-domain of lactoferrin is sufficient for binding, internalization and targeting to the nucleus of host cells. The capacity of blactoferrin to reach the nucleus has been shown in different cell monolayers, including intestinal cells 73, and in freshly-isolated monocytes 68. The blactoferrin ability to localize into the nuclei of these cells is comparable to that showed by hlactoferrin in endothelial cells 74. Consequently, the nuclear localization strongly suggested that this molecule may be involved in the transcriptional regulation of some genes of host inflammatory responses, thus acting as a transcriptional factor and modulator of the inflammatory processes through the inhibition of pro-inflammatory cytokines 75.
Lactoferrin and Anemia of Inflammation
Although the mechanisms by which bovine lactoferrin exerts its anti-inflammatory activity are under debate, in 2006, by designing the first clinical trial on the effect of 30 days of bovine lactoferrin oral administration (100 mg two times a day before meals) in pregnant women with iron deficiency anemia or anemia of inflammation, a group of scientists obtained surprising results 76. In fact, pregnant women receiving 100 mg of bovine lactoferrin, iron saturated at 20–30% two times a day acquired 70–84 µg/day of iron, respectively. Although the concentration of iron supplemented by bovine lactoferrin is very far from that which is required daily (1–2 mg), a significant increase of the concentration of hemoglobin (Hb) and total serum iron was detected after 30 days of the treatment. Therefore, researchers speculated that bovine lactoferrin efficacy in curing anemia of inflammation was presumably not linked to direct iron supplementation, but to a more complex mechanism involving this protein in iron homeostasis. Later on, in other clinical trials, bovine lactoferrin treatment showed a significant improvement of hematological parameters, including red blood cell number, hemoglobin, total serum iron, serum ferritin concentrations and percentage of hematocrit, in pregnant women suffering from iron deficiency anemia, associated with a consistent decrease of serum IL-6 levels 73. Lactoferrin is, therefore, a key element, not only in the host defense system 77, but also a pivotal component able to inhibit the inflammatory response, especially in inflamed pregnant women affected by hereditary thrombophilia 78.
- Lactoferrin: A Natural Glycoprotein Involved in Iron and Inflammatory Homeostasis. Int. J. Mol. Sci. 2017, 18(9), 1985; doi:10.3390/ijms18091985 https://www.mdpi.com/1422-0067/18/9/1985/htm[↩][↩][↩][↩][↩]
- Johansson, B. Isolation of an iron-containing red protein from human milk. Acta Chem. Scand. 1960, 14, 510–512.[↩]
- Groves, M.L. The isolation of a red protein from milk. J. Am. Chem. Soc. 1960, 82, 3345–3350.[↩]
- Masson, P.L.; Heremans, J.F. Lactoferrin in milk from different species. Comp. Biochem. Physiol. 1971, B39, 119–129.[↩]
- Masson, P.L.; Heremans, J.F.; Schonne, E. Lactoferrin, an iron-binding protein in neutrophilic leukocytes. J. Exp. Med. 1969, 130, 643–658.[↩]
- Anderson, B.F.; Baker, H.M.; Norris, G.E.; Rice, D.W.; Baker, E.N. Structure of human lactoferrin: Crystallographic structure analysis and refinement at 2.8 A resolution. J. Mol. Biol. 1989, 209, 711–734.[↩][↩]
- Gerstein, M.; Anderson, B.F.; Norris, G.E.; Baker, E.N.; Lesk, A.M.; Chothia, C. Domain closure in lactoferrin. Two hinges produce a see-saw motion between alternative close-packed interfaces. J. Mol. Biol. 1993, 234, 357–372.[↩]
- Baker, H.M.; Baker, E.N. A structural perspective on lactoferrin function. Biochem. Cell Biol. 2012, 90, 320–328.[↩]
- Kumar, H.; Wen, J.; Shaw, J.; Cornish, J.; Bunt, C. Physiochemical characterization of β-glucan and in vitro release of lactoferrin from β-glucan microparticles. Curr. Drug Deliv. 2013, 10, 713–721.[↩]
- Woodman T, Strunk T, Patole S, Hartmann B, Simmer K, Currie A. Effects of lactoferrin on neonatal pathogens and Bifidobacterium breve in human breast milk. PLoS One. 2018;13(8):e0201819. Published 2018 Aug 22. doi:10.1371/journal.pone.0201819 https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6104981/[↩]
- Ellison RT, Giehl TJ, LaForce FM. Damage of the outer membrane of enteric gram-negative bacteria by lactoferrin and transferrin. Infection and Immunity. 1988;56(11):2774–81[↩]
- Drago-Serrano ME, Campos-Rodriguez R, Carrero JC, de la Garza M. Lactoferrin: Balancing Ups and Downs of Inflammation Due to Microbial Infections. Int J Mol Sci. 2017;18(3). Epub 2017/03/04. 10.3390/ijms18030501[↩]
- Embleton ND, Berrington JE, Dorling J, Ewer AK, Juszczak E, Kirby JA, et al. Mechanisms Affecting the Gut of Preterm Infants in Enteral Feeding Trials. Frontiers in Nutrition. 2017;4:14 10.3389/fnut.2017.00014[↩][↩]
- Born too soon: the global epidemiology of 15 million preterm births. Blencowe H, Cousens S, Chou D, Oestergaard M, Say L, Moller AB, Kinney M, Lawn J, Born Too Soon Preterm Birth Action Group. Reprod Health. 2013; 10 Suppl 1():S2[↩]
- Distortions in development of intestinal microbiota associated with late onset sepsis in preterm infants. Mai V, Torrazza RM, Ukhanova M, Wang X, Sun Y, Li N, Shuster J, Sharma R, Hudak ML, Neu J PLoS One. 2013; 8(1):e52876.[↩]
- Intestinal colonization patterns of staphylococci in preterm infants in relation to type of enteral feeding and bacteremia. Cossey V, Vanhole C, Verhaegen J, Schuermans A. Breastfeed Med. 2014 Mar; 9(2):79-85.[↩]
- The preterm gut microbiota: changes associated with necrotizing enterocolitis and infection. Stewart CJ, Marrs EC, Magorrian S, Nelson A, Lanyon C, Perry JD, Embleton ND, Cummings SP, Berrington JE. Acta Paediatr. 2012 Nov; 101(11):1121-7.[↩]
- Moles L, Manzano S, Fernández L, Montilla A, Corzo N, Ares S, et al. Bacteriological, biochemical, and immunological properties of colostrum and mature milk from mothers of extremely preterm infants. Journal of Pediatric Gastroenterology and Nutrition. 2015;60(1):120–6. 10.1097/MPG.0000000000000560 https://www.ncbi.nlm.nih.gov/pubmed/25207476[↩]
- Trend S, Strunk T, Hibbert J, Kok CH, Zhang G, Doherty DA, et al. Antimicrobial protein and peptide concentrations and activity in human breast milk consumed by preterm infants at risk of late-onset neonatal sepsis. PLoS ONE. 2015;10(2):0117038[↩]
- Jakaitis BM, Denning PW. Commensal and probiotic bacteria may prevent NEC by maturing intestinal host defenses. Pathophysiology. 2014;21(1):47–54. Epub 2014/01/21. 10.1016/j.pathophys.2013.11.012[↩]
- Patole SK, Rao SC, Keil AD, Nathan EA, Doherty DA, Simmer KN. Benefits of Bifidobacterium breve M-16V Supplementation in Preterm Neonates—A Retrospective Cohort Study. PLoS ONE. 2016;11(3). 10.1371/journal.pone.0150775[↩]
- Manzoni P, Rinaldi M, Cattani S, et al. Bovine lactoferrin supplementation for prevention of late-onset sepsis in very low-birth-weight neonates: a randomized trial. The Journal of the American Medical Association. 2009;302(13):1421–8. 10.1001/jama.2009.1403[↩]
- Puddu, P.; Latorre, D.; Carollo, M.; Catizone, A.; Ricci, G.; Valenti, P.; Gessani, S. Bovine lactoferrin counteracts Toll-like receptor mediated activation signals in antigen presenting cells. PLoS ONE 2011, 6, e22504[↩]
- Weinberg, E.D. The development of awareness of iron-withholding defense. Perspect. Biol. Med. 1993, 36, 215–221[↩]
- Petrik, M.; Zhai, C.; Haas, H.; Decristoforo, C. Siderophores for molecular imaging applications. Clin. Transl. Imaging 2017, 5, 15–27.[↩]
- Pogoutse, A.K.; Moraes, T.F. Iron acquisition through the bacterial transferrin receptor. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 314–326.[↩]
- Wandersman, C.; Stojiljkovic, I. Bacterial heme sources: The role of heme, hemoprotein receptors and hemophores. Curr. Opin. Microbiol. 2000, 3, 215–220.[↩]
- Huang, W.; Wilks, A. Extracellular Heme Uptake and the Challenge of Bacterial Cell Membranes. Annu. Rev. Biochem. 2017, 86, 799–823.[↩]
- Pilarczyk-Zurek, M.; Strus, M.; Adamski, P.; Heczko, P.B. The dual role of Escherichia coli in the course of ulcerative colitis. BMC Gastroenterol. 2016, 16, 128.[↩]
- Singh, P.K.; Parsek, M.R.; Greenberg, E.P.; Welsh, M.J. A component of innate immunity prevents bacterial biofilms development. Nature 2002, 417, 552–555.[↩][↩]
- Stites, S.W.; Walters, B.; O’Brien-Ladner, A.R.; Bailey, K.; Wesselius, L.J. Increased iron and ferritin content of sputum from patients with cystic fibrosis or chronic bronchitis. Chest 1998, 114, 814–819.[↩]
- Berlutti, F.; Morea, C.; Battistoni, A.; Sarli, S.; Cipriani, P.; Superti, F.; Ammendolia, M.G.; Valenti, P. Iron availability influences aggregation, biofilm, adhesion and invasion of Pseudomonas aeruginosa and Burkholderia cenocepacia. Int. J. Immunopathol. Pharmacol. 2005, 18, 661–670.[↩]
- Rogan, M.P.; Taggart, C.C.; Greene, C.M.; Murphy, P.G.; O’Neill, S.J.; McElvaney, N.G. Loss of microbicidal activity and increased formation of biofilm due to decreased lactoferrin activity in patients with cystic fibrosis. J. Infect. Dis. 2004, 190, 1245–1253.[↩]
- Thompson, A.B.; Bohling, T.; Payvandi, F.; Rennard, S.I. Lower respiratory tract lactoferrin and lysozyme arise primarily in the airways and are elevated in association with chronic bronchitis. J. Lab. Clin. Med. 1990, 115, 148–158.[↩]
- Brandenburg, K.; Jurgens, G.; Muller, M.; Fukuoka, S.; Koch, M.H.J. Biophysical characterization of lipopolysaccharide and lipid A inactivation by lactoferrin. Biol. Chem. 2001, 382, 1215–1225.[↩]
- Bellamy, W.; Takase, M.; Wakabayashi, H.; Kavase, K.; Tomita, M. Antibacterial spectrum of lactoferricin B, a potent bactericide peptide derived from the N-terminal region of bovine lactoferrin. J. Appl. Bacteriol. 1992, 73, 472–479.[↩]
- Rossi, P.; Giansanti, F.; Boffi, A.; Ajello, M.; Valenti, P.; Chiancone, E.; Antonini, G. Ca2+ binding to bovine lactoferrin enhances protein stability and influences the release of bacterial lipopolysaccharide. Biochem. Cell Biol. 2002, 80, 41–48.[↩]
- Valenti, P.; Antonini, G. Lactoferrin: An important host defence against microbial and viral attack. Cell. Mol. Life Sci. 2005, 62, 2576–2587.[↩][↩]
- Visca, P.; Berlutti, F.; Vittorioso, P.; Dalmastri, C.; Thaller, M.C.; Valenti, P. Growth and adsorption of Streptococcus mutans 6715–13 to hydroxyapatite in the presence of lactoferrin. Med. Microbiol. Immunol. 1989, 178, 69–79.[↩]
- Oho, T.; Mitoma, M.; Koga, T. Functional domain of bovine milk lactoferrin which inhibits the adherence of Streptococcus mutans cells to a salivary film. Infect. Immun. 2002, 70, 5279–5282.[↩]
- Williams, T.J.; Schneider, R.P.; Willcox, M.D. The effect of protein-coated contact lenses on the adhesion and viability of gram negative bacteria. Curr. Eye Res. 2003, 27, 227–235.[↩]
- Longhi, C.; Conte, M.P.; Seganti, L.; Polidoro, M.; Alfsen, A.; Valenti, P. Influence of lactoferrin on the entry process of Escherichia coli HB101 (pRI203) in HeLa cells. Med. Microbiol. Immunol. 1993, 182, 25–35.[↩][↩][↩]
- Kawasaki, Y.; Tazume, S.; Shimizu, K.; Matsuzawa, H.; Dosako, S.; Isoda, H.; Tsukiji, M.; Fujimura, R.; Muranaka, Y.; Isihida, H. Inhibitory effects of bovine lactoferrin on the adherence of enterotoxigenic Escherichia coli to host cells. Biosci. Biotechnol. Biochem. 2000, 64, 348–354.[↩][↩]
- Ammendolia, M.G.; Bertuccini, L.; Iosi, F.; Minelli, F.; Berlutti, F.; Valenti, P.; Superti, F. Bovine lactoferrin interacts with cable pili of Burkholderia cenocepacia. Biometals 2010, 23, 531–542.[↩]
- Araújo, A.N.; Giugliano, L.G. Lactoferrin and free secretory component of human milk inhibit the adhesion of enteropathogenic Escherichia coli to HeLa cells. BMC Microbiol. 2001, 1, 25.[↩][↩]
- Dalmastri, C.; Valenti, P.; Visca, P.; Vittorioso, P.; Orsi, N. Enhanced antimicrobial activity of lactoferrin by binding to the bacterial surface. Microbiologica 1988, 11, 225–230.[↩]
- Wu, H.F.; Monroe, D.M.; Church, F.C. Characterization of the glycosaminoglycan-binding region of Lactoferrin. Arch. Biochem. Biophys. 1995, 317, 85–92.[↩][↩]
- Shimazaki, K.; Tazume, T.; Uji, K.; Tanaka, M.; Kumura, H.; Mikawa, K.; Shimo-Oka, T. Properties of a heparin-binding peptide derived from bovine lactoferrin. J. Dairy Sci. 1998, 81, 2841–2849.[↩]
- Willer Eda, M.; Lima Rde, L.; Giugliano, L.G. In vitro adhesion and invasion inhibition of Shigella dysenteriae, Shigella flexneri and Shigella sonnei clinical strains by human milk proteins. BMC Microbiol. 2004, 4, 18–25.[↩]
- Gomez, H.F.; Ochoa, T.J.; Carlin, L.G.; Cleary, T.G. Human lactoferrin impairs virulence of Shigella flexneri. J. Infect. Dis. 2003, 187, 87–95.[↩]
- Dial, E.J.; Lichtenberger, L.M. Effect of lactoferrin on Helicobacter felis induced gastritis. Biochem. Cell Biol. 2002, 80, 113–117[↩]
- Qiu, J.; Hendrixson, D.R.; Baker, E.N.; Murphy, T.F.; St Geme, J.W.; Plaut, A.G. Human milk lactoferrin inactivates two putative colonization factors expressed by Haemophilus influenzae. Proc. Natl. Acad. Sci. USA 1998, 95, 12641–12646.[↩]
- Ochoa, T.J.; Noguera-Obenza, M.; Ebel, F.; Guzman, C.A.; Gomez, H.F.; Cleary, T.G. Lactoferrin impairs type III secretory system function in enteropathogenic Escherichia coli. Infect. Immun. 2003, 71, 5149–5155.[↩]
- Hendrixson, D.R.; Qiu, J.; Shewry, S.C.; Fink, D.L.; Petty, S.; Baker, E.N.; Plaut, A.G.; St. Geme, J.W. Human milk lactoferrin is a serine protease that cleaves Haemophilus surface proteins at arginine-rich sites. Mol. Microbiol. 2003, 47, 607–617.[↩]
- Berlutti, F.; Schippa, S.; Morea, C.; Sarli, S.; Perfetto, B.; Donnarumma, G.; Valenti, P. Lactoferrin downregulates pro-inflammatory cytokines up-expressed in intestinal epithelial cells infected with invasive or noninvasive Escherichia coli strains. Biochem. Cell Biol. 2006, 84, 351–357.[↩][↩][↩]
- Armstead, A.L.; Li, B. Nanomedicine as an emerging approach against intracellular pathogens. Int. J. Nanomed. 2011, 6, 3281–3293.[↩]
- Di Biase, A.M.; Tinari, A.; Pietrantoni, A.; Antonini, G.; Valenti, P.; Conte, M.P.; Superti, F. Effect of bovine lactoferricin on enteropathogenic Yersinia adhesion and invasion in HEp-2 cells. J. Med. Microbiol. 2004, 53, 407–412.[↩]
- Ochoa, T.J.; Brown, E.L.; Guion, C.E.; Chen, J.Z.; McMahon, R.J.; Cleary, T.G. Effect of lactoferrin on enteroaggregative E. coli (EAEC) 1. Biochem. Cell Biol. 2006, 84, 369–376.[↩]
- Ajello, M.; Greco, R.; Giansanti, F.; Massucci, M.T.; Antonini, G.; Valenti, P. Anti-invasive activity of bovine lactoferrin towards group A streptococci. Biochem. Cell Biol. 2002, 80, 119–124.[↩][↩]
- Valenti, P.; Antonini, G. Lactoferrin: An important host defence against microbial and viral attack. Cell. Mol. Life Sci. 2005, 62, 2576–2587[↩]
- Frioni, A.; Conte, M.P.; Cutone, A.; Longhi, C.; Musci, G.; di Patti, M.C.; Natalizi, T.; Marazzato, M.; Lepanto, M.S.; Puddu, P.; et al. Lactoferrin differently modulates the inflammatory response in epithelial models mimicking human inflammatory and infectious diseases. Biometals 2014, 27, 843–856.[↩][↩]
- Sessa, R.; Di Pietro, M.; Filardo, S.; Bressan, A.; Rosa, L.; Cutone, A.; Frioni, A.; Berlutti, F.; Paesano, R.; Valenti, P. Effect of bovine lactoferrin on Chlamydia trachomatis infection and inflammation. Biochem. Cell Biol. 2017, 95, 34–40.[↩][↩][↩][↩][↩][↩]
- Kohlhoff, S.A.; Hammerschlag, M.R. Treatment of Chlamydial infections: 2014 update. Expert Opin. Pharmacother. 2015, 16, 205–212.[↩]
- Stallmann, S.; Hegemann, J.H. The Chlamydia trachomatis Ctad1 invasin exploits the human integrin 1 receptor for host cell entry. Cell. Microbiol. 2015, 18, 761–775.[↩]
- Lu, L.; Hangoc, G.; Oliff, A.; Chen, L.T.; Shen, R.N.; Broxmeyer, H.E. Protective influence of lactoferrin on mice infected with the polycythemia-inducing strain of Friend virus complex. Cancer Res. 1987, 47, 4184–4188.[↩]
- Berlutti, F.; Pantanella, F.; Natalizi, T.; Frioni, A.; Paesano, R.; Polimeni, A.; Valenti, P. Antiviral properties of lactoferrin—A natural immunity molecule. Molecules 2011, 16, 6992–7018.[↩]
- Osterlund, P.J.; Pirhonen, N.; Ikonen, E.; Ronkko, M.; Strengell, S.M.; Makela, M.; Broman, O.J.; Hamming, R.; Hartmann, T.; Ziegler, T.; et al. Pandemic H1N1 2009 influenza A virus induces weak cytokine responses in human macrophages and dendritic cells and is highly sensitive to the antiviral actions of interferons. J. Virol. 2010, 84, 1414–1422.[↩]
- Puddu, P.; Latorre, D.; Carollo, M.; Catizone, A.; Ricci, G.; Valenti, P.; Gessani, S. Bovine lactoferrin counteracts Toll-like receptor mediated activation signals in antigen presenting cells. PLoS ONE 2011, 6, e22504.[↩][↩]
- Latorre, D.; Berlutti, F.; Valenti, P.; Gessani, S.; Puddu, P. LF immunomodulatory strategies: Mastering bacterial endotoxin. Biochem. Cell Biol. 2012, 90, 269–278.[↩]
- Vora, P.; Youdim, A.; Thomas, L.S.; Fukata, M.; Tesfay, S.Y.; Lukasek, K.; Michelsen, K.S.; Wada, A.; Hirayama, T.; Arditi, M.; et al. Beta-defensin-2 expression is regulated by TLR signaling in intestinal epithelial cells. J. Immunol. 2004, 173, 5398–5405.[↩]
- Ashida, K.; Sasaki, H.; Suzuki, Y.A.; Lönnerdal, B. Cellular internalization of lactoferrin in intestinal epithelial cells. Biometals 2004, 17, 311–315[↩]
- Suzuki, Y.A.; Wong, H.; Ashida, K.Y.; Schryvers, A.B.; Lönnerdal, B. The N1 domain of human lactoferrin is required for internalization by caco-2 cells and targeting to the nucleus. Biochemistry 2008, 47, 10915–10920.[↩]
- Paesano, R.; Natalizi, T.; Berlutti, F.; Valenti, P. Body iron delocalization: The serious drawback in iron disorders in both developing and developed countries. Pathog. Glob. Health 2012, 106, 200–216.[↩][↩]
- Kim, C.W.; Lee, T.H.; Park, K.H.; Choi, S.Y.; Kim, J. Human lactoferrin suppresses TNF-a-induced intercellular adhesion molecule-1 expression via competition with NF-kB in endothelial cells. FEBS Lett. 2012, 586, 229–234.[↩]
- Paesano, R.; Pietropaoli, M.; Gessani, S.; Valenti, P. The influence of lactoferrin, orally administered, on systemic iron homeostasis in pregnant women suffering of iron deficiency and iron deficiency anaemia. Biochimie 2009, 91, 44–51.[↩]
- Paesano, R.; Torcia, F.; Berlutti, F.; Pacifici, E.; Ebano, V.; Moscarini, M.; Valenti, P. Oral administration of lactoferrin increases hemoglobin and total serum iron in pregnant women. Biochem. Cell Biol. 2006, 84, 377–380.[↩]
- Legrand, D.; Elass, E.; Carpentier, M.; Mazurier, J. Lactoferrin: A modulator of immune and inflammatory responses. Cell. Mol. Life Sci. 2005, 62, 2549–2559.[↩]
- Paesano, R.; Pacifici, E.; Benedetti, S.; Berlutti, F.; Frioni, A.; Polimeni, A.; Valenti, P. Safety and efficacy of lactoferrin versus ferrous sulphate in curing iron deficiency and iron deficiency anaemia in hereditary thrombophilia pregnant women: An interventional study. Biometals 2014, 27, 999–1006.[↩]

{kind=link}