Hemagglutinin
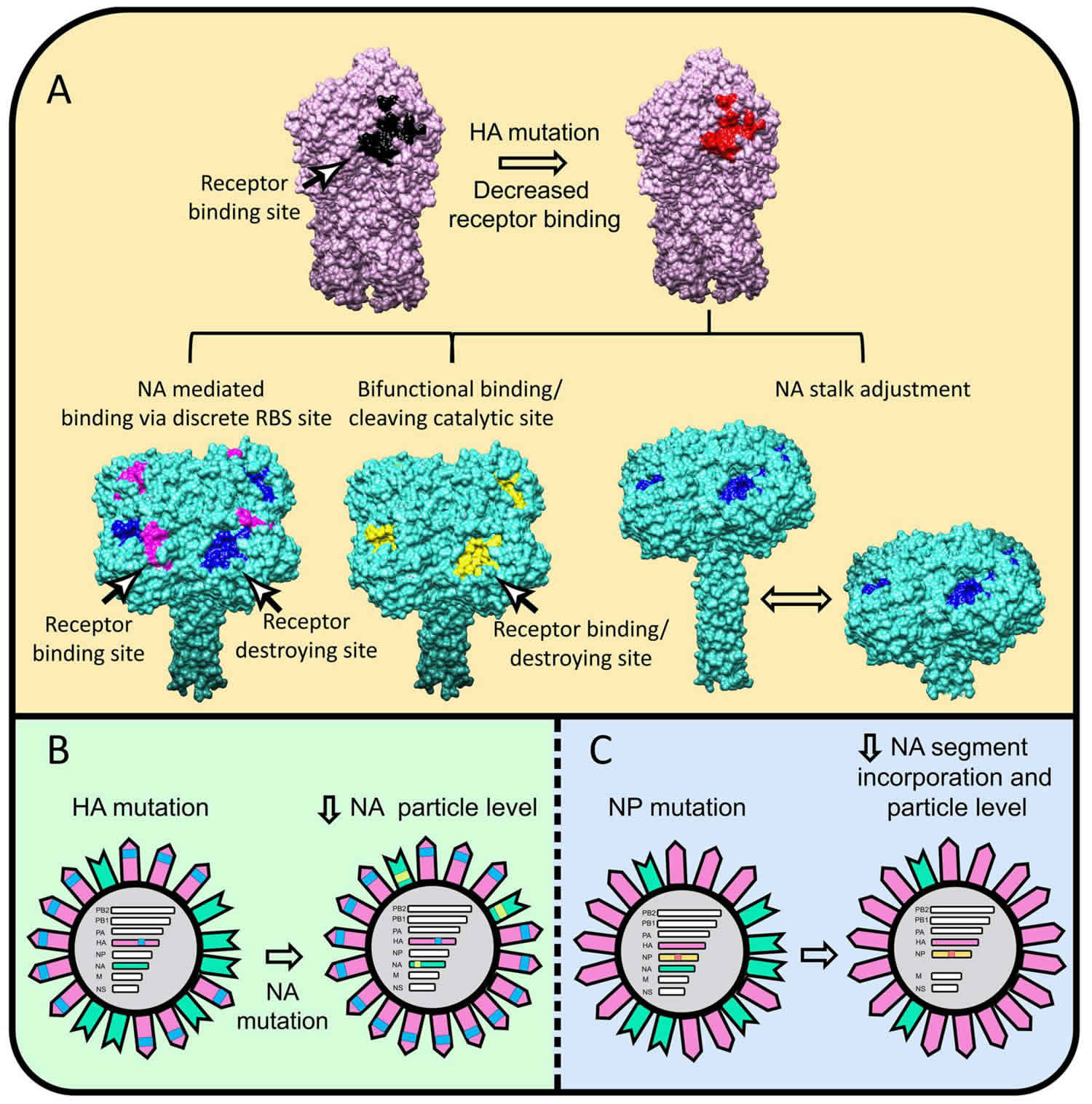
Hemagglutinin (HA) is a viral glycan-binding protein or an antigenic glycoprotein found on the surface of the influenza viruses (as well as many other bacteria and viruses). Hemagglutinin is responsible for binding the virus to the cell that is being infected. The name “hemagglutinin” comes from the protein’s ability to cause red blood cells (erythrocytes) to clump together (“agglutinate”) in vitro 1. The surface glycoproteins hemagglutinin (HA) and neuraminidase (NA) are how the H1N1-Swine flu (a subtype of influenza A virus) is differentiated from other strains of influenza A (H1N1, H1N2) depending on the type of hemagglutinin (HA) or neuraminidase (NA) antigens expressed with metabolic synergy 2. The function of hemagglutinin (HA) is to cause red blood cells to cluster together, and it attaches the virus to the infected cell by binding to terminal sialic acid residues on glycoproteins/glycolipids to initiate the infectious cycle 3. Neuraminidase (NA) helps move the virus particles through the infected cell and assists in budding from host cells by cleaving terminal sialic acids, releasing the virus to complete the infectious cycle 3. The hemagglutinin is also the major antigen against which neutralizing antibodies are produced, and antigenic changes in this protein are in part responsible for new viral outbreaks and considered in formulation of the annual influenza vaccine.
There are at least 16 different hemagglutinin (HA) antigens. These subtypes are labeled H1 through H16. The last, H16, was discovered only recently on influenza A viruses isolated from black-headed gulls from Sweden and Norway 4. The first three hemagglutinins, H1, H2, and H3, are found in human influenza viruses.
A highly pathogenic avian flu virus of H5N1 type has been found to infect humans at a low rate. It has been reported that single amino acid changes in this avian virus strain’s type H5 hemagglutinin have been found in human patients that “can significantly alter receptor specificity of avian H5N1 viruses, providing them with an ability to bind to receptors optimal for human influenza viruses” 5. This finding seems to explain how an H5N1 virus that normally does not infect humans can mutate and become able to efficiently infect human cells. The hemagglutinin of the H5N1 virus has been associated with the high pathogenicity of this flu virus strain, apparently due to its ease of conversion to an active form by proteolysis 6.
Influenza virus hemagglutinin
The most well-studied example of a viral glycan-binding protein is the influenza virus hemagglutinin (HA), which binds to sialic acid–containing glycans on the host cell surface. Although the affinity of this interaction is low, like that of other lectins with their glycan ligands, the avidity is increased by hemagglutinin trimerization and a high density of glycan receptors present on the host cell. This binding event is required for internalization of the virus by endocytosis and subsequently the pH-dependent fusion of the viral envelope with the endosomal membrane, ultimately triggering release of the viral RNA into the cytosol. The specificity of the host glycan-hemagglutinin interaction varies considerably for different subtypes of influenza. For example, human strains of influenza-A and -B viruses bind primarily to cells containing N-acetylneuraminic acid (Neu5Acα)2–6Gal-containing receptors. However, avian influenza viruses bind to receptors expressing Neu5Acα2–3Gal-, and porcine strains bind to both Neu5Acα2–6Gal- and -3Gal-containing receptors. This linkage preference is a result of structural differences of the hemagglutinin.
Influenza viruses express two types of antigens; hemagglutinin (HA) and neuraminidase (NA). Influenza A virus has 18 hemagglutinin (HA), and 11 neuraminidase (NA) subtypes and these antigens are critical for the virulence of the organism 7. The trimeric hemagglutinin glycoprotein acts by promoting attachment of the virus to the host cell surface resulting in fusion and thereby releasing virions into the cytoplasm 8.
Differently combined hemagglutinin (HA) and neuraminidase (NA) antigens are seen in influenza A, that in turn undergo antigenic drifts and shifts resulting in antigenic variation and thereby the necessity for vaccine strain types to vary accordingly. Antigenic drifts are genetic changes occurring in the virus due to various actions of polymerases leading to gradual antigenic changes in both hemagglutinin and neuraminidase producing new variant strains. An antigenic shift takes place when the currently circulating virus disappears and gets replaced by a new subtype with novel glycoproteins to which antibodies against the previously circulating subtype do not cross-react 9.
Influenza vaccine conveys immunity against the influenza virus by stimulating the production of antibodies specific to the disease. Antibodies to neuraminidase act by aggregating viruses on the cell surface effectively and reducing the amount of virus released from infected cells. Regarding the induction of immunity, the surface hemagglutinin protein of the influenza virus contains two structural elements, head and stalk wherein the head is the primary target of antibodies that confer protective immunity against influenza viruses 8
Flu shots offer protection against three or four strains of the flu virus. Trivalent flu vaccines provide protection against two influenza A strains, H1N1 and H3N2, and one influenza B strain. Quadrivalent flu vaccines protect against the same strains as the trivalent vaccine as well as an additional strain of influenza B 10.
The mechanism of immune protection is more complicated, as while primarily humoral, cell-mediated immunity also plays an essential role in immunity to influenza. After vaccination, it takes two weeks to build up an immune response against the flu. The effectiveness of a vaccine depends on several host factors such as age, underlying health status, genetic status and furthermore on antigenic matches between the vaccine and circulating viruses 11.
References- Nelson DL and Cox MM, 2005. Lehninger’s Principles of Biochemistry, 4th edition, WH Freeman, New York, NY.
- Jilani TN, Jamil RT, Siddiqui AH. H1N1 Influenza (Swine Flu) [Updated 2019 Jun 9]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2019 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK513241
- Kosik, I.; Yewdell, J.W. Influenza Hemagglutinin and Neuraminidase: Yin–Yang Proteins Coevolving to Thwart Immunity. Viruses 2019, 11, 346. https://www.mdpi.com/1999-4915/11/4/346
- Suzuki, Y, 2005. Sialobiology of Influenza: Molecular Mechanism of Host Range Variation of Influenza Viruses in Biological and Pharmaceutical Bulletin, vol 28, pp399-408
- Gambaryan A, Tuzikov A, Pazynina G, Bovin N, Balish A, Klimov A, 2006. Evolution of the receptor binding phenotype of influenza A (H5) viruses in Virology vol 344, issue 2, pp432-8
- Hatta M, Gao P, Halfmann P, Kawaoka Y, 2001. Molecular Basis for High Virulence of Hong Kong H5N1 Influenza A Viruses in Science vol 293, pp1840-1842.
- Kalarikkal SM, Jaishankar GB. Influenza Vaccine. [Updated 2019 Oct 8]. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2019 Jan-. Available from: https://www.ncbi.nlm.nih.gov/books/NBK537197
- Kirkpatrick E, Qiu X, Wilson PC, Bahl J, Krammer F. The influenza virus hemagglutinin head evolves faster than the stalk domain. Sci Rep. 2018 Jul 11;8(1):10432.
- Franco-Paredes C, Carrasco P, Preciado JI. The first influenza pandemic in the new millennium: lessons learned hitherto for current control efforts and overall pandemic preparedness. J Immune Based Ther Vaccines. 2009 Aug 07;7:2.
- Rudenko L, Kiseleva I, Krutikova E, Stepanova E, Rekstin A, Donina S, Pisareva M, Grigorieva E, Kryshen K, Muzhikyan A, Makarova M, Sparrow EG, Torelli G, Kieny MP. Rationale for vaccination with trivalent or quadrivalent live attenuated influenza vaccines: Protective vaccine efficacy in the ferret model. PLoS ONE. 2018;13(12):e0208028
- Lim JW, Na W, Kim HO, Yeom M, Park G, Kang A, Chun H, Park C, Oh S, Le VP, Jeong HH, Song D, Haam S. Cationic Poly(Amino Acid) Vaccine Adjuvant for Promoting Both Cell-Mediated and Humoral Immunity Against Influenza Virus. Adv Healthc Mater. 2019 Jan;8(2):e1800953



{kind=link}