
What is quercetin
Quercetin is a polyphenolic flavonoid widely distributed in many plants and fruits including red grapes, citrus fruit, tomato, broccoli and other leafy green vegetables, and a number of berries, including raspberries and cranberries 1. Quercetin acutally belongs to a type of flavonols, which is a class of flavonoids that have the 3-hydroxyflavone backbone (3-hydroxy-2-phenylchromen-4-one) and present in a wide variety of herbs including licorice 2. Quercetin is a licorice flavonoid and its IUPAC name is 2-(3,4-dihydroxyphenyl)-3,5,7-trihydroxy-4H-chromen-4-one.
Several studies demonstrated that quercetin had diverse bioactive effects such as antiviral activity, anti-asthmatic activity, anti-oxidant, monoamine-oxidase inhibitor, anti-tumor and anti-inflammatory activity and quercetin has been studied extensively as a chemoprevention agent in several cancer models 3, 4, 5. Quercetin has been used in medicine to decrease capillary fragility. It has also been used in dyes and as a veterinary drug 6. However, there is no reliable clinical evidence that quercetin has a protective effect on human cancer.
Quercetin itself (aglycone quercetin), as opposed to quercetin glycosides, is not a normal dietary component. Quercitin glycosides are converted to phenolic acids as they pass through the gastrointestinal tract. Quercetin has neither been confirmed scientifically as a specific therapeutic for any condition nor been approved by any regulatory agency. The U. S. Food and Drug Administration has not approved any health claims for quercetin. Nevertheless, the interest in dietary flavonoids has grown after the publication of several epidemiological studies showing an inverse correlation between dietary consumption of flavonols and flavones and reduced incidence and mortality from cardiovascular disease and cancer. In recent years, a large amount of experimental and some clinical data have accumulated regarding the effects of flavonoids on the endothelium under physiological and pathological conditions. The meta-analysis of seven prospective cohort studies concluded that the individuals in the top third of dietary flavonol intake are associated with a reduced risk of mortality from coronary heart disease as compared with those in the bottom third, after adjustment for known risk factors and other dietary components. A limited number of intervention studies with flavonoids and flavonoid containing foods and extracts has been performed in several pathological conditions.
Flavonoids are a large group of compounds (>4000) that occur naturally in fruits, vegetables, nuts, seeds, flowers and other plant matter and as such they are an integral part of the human diet 7. They all share a common three-ring structure but are subdivided into flavonols, flavons, flavanols and flavanons according to their substituents 8. Epidemiological studies indicate that diets rich in flavonoids are associated with reduced incidences of several chronic diseases including cardiovascular disease, asthma, type II diabetes and certain types of cancer 9. The cardioprotective properties of flavonoids are multi-faceted involving antioxidant, anti-hypercholesterolaemia, anti-inflammatory and inhibition of platelet aggregation effects 9. The antioxidant property of flavonoids was thought, until relatively recently, to underlie the majority of their protective cellular effects. However, it is becoming increasingly apparent that flavonoids also influence cellular function by modulating the activity of many enzymes including the inhibition of protein kinases and lipid kinases 10.
The inhibitory effect of flavonoids on protein kinase activity is primarily due to their ability to function as competitive inhibitors of the ATP-binding domain located in the active site of these enzymes 10. Protein kinases directly inhibited by flavonoids include protein kinase C (PKC), myosin light chain kinase (MLCK) and MEK1 10. MEK1 is an upstream kinase responsible for the activation of extracellular signal-regulated kinase 1/2 (ERK1/2), a member of the mitogen-activated protein kinase (MAPK) family of protein kinases 11. This family also includes p38 MAPK and JNK signalling pathways, both of which are targets for modulation by flavonoids 12. It is also notable that flavonoids such as quercetin also inhibit (via competitive inhibition of the ATP-binding site) the lipid kinase PI-3Kγ 13, which plays a prominent role in the activation of protein kinase B (PKB; also known as Akt).
The MEK/ERK1/2 and the PI-3K/PKB pathways are important signalling pathways which mediate cell survival and cardioprotection against ischaemia/reperfusion injury 14. In contrast, p38 MAPK and JNK cascades are typically associated with inflammation and cell death; however, there is also evidence that they mediate cardioprotection 14. Hence, the cardioprotective effects of flavonoids may relate to their ability to modulate MAPK and PI-3K/PKB signalling cascades. Indeed, recent studies have shown that grape seed proanthocyanidin extract protects cardiomyocytes from ischaemia/reperfusion injury via PKB activation 15. Similarly, quercetin (the most abundant dietary flavonoid) protected H9c2 cardiomyoblasts from H2O2-induced cell death via enhancement of ERK1/2 and PI-3K/PKB signalling 16. In contrast, isorhamnetin (a flavonol isolated from sea buckthorn) prevented H2O2-induced activation of apoptosis in H9c2 cells through ERK1/2 inactivation 17.
Figure 1. Quercetin
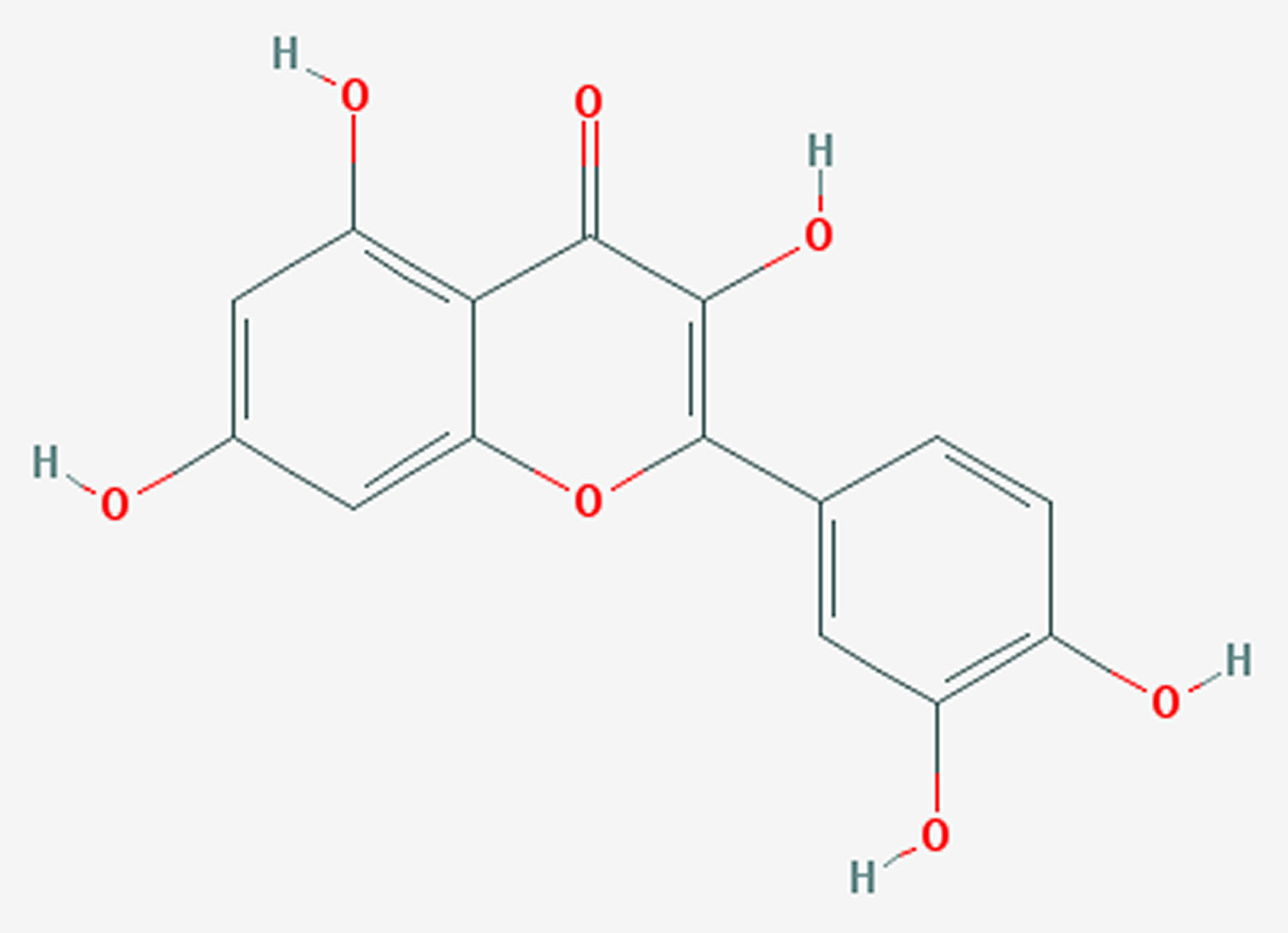
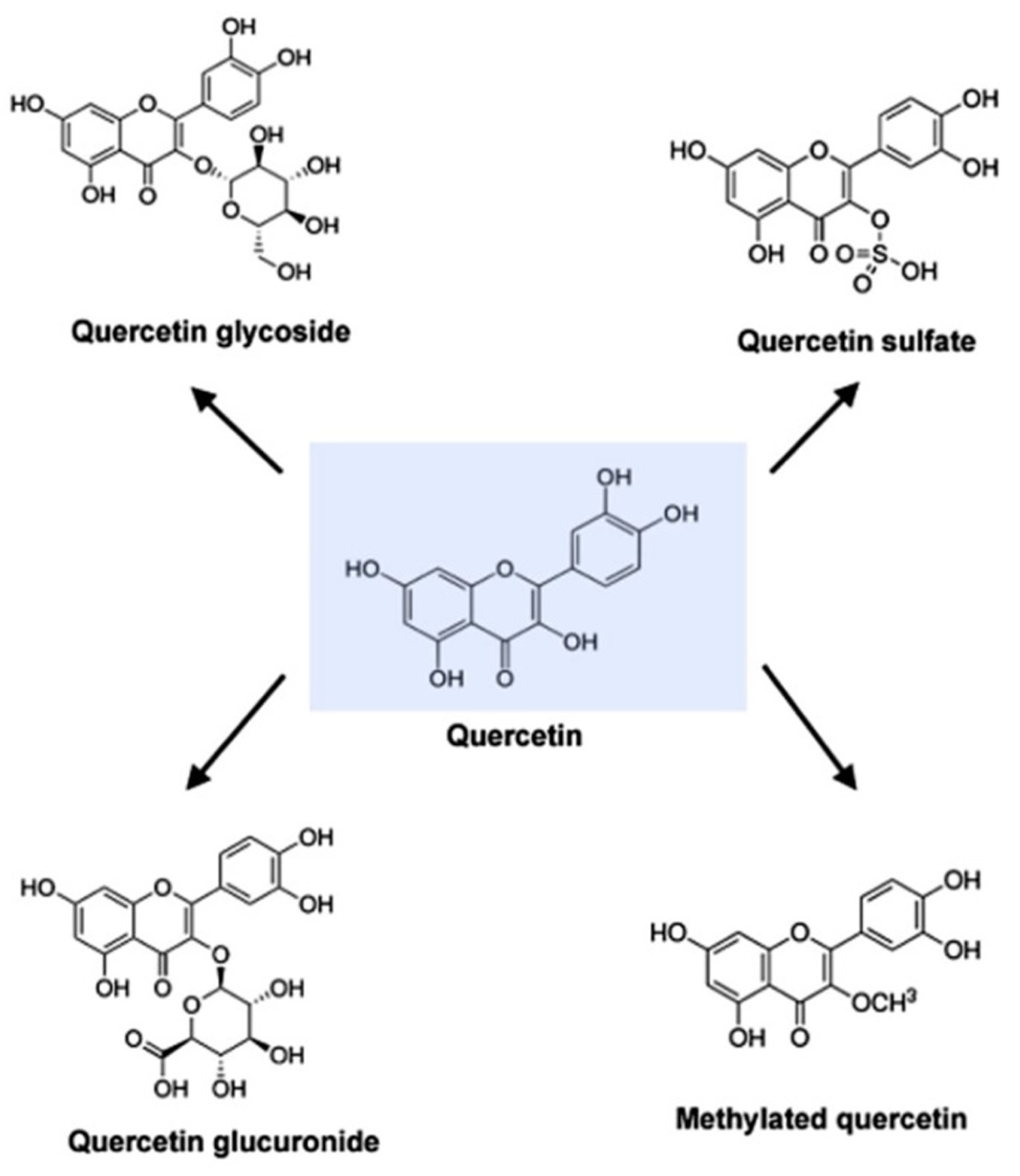
Figure 2. Molecular structure of quercetin, quercetin glycoside, quercetin glucuronide, quercetin sulfate and methylated quercetin
Note:
Quercetin (C15H10O7) is an aglycone, lacking an attached sugar. It is a brilliant citron yellow needle crystal and entirely insoluble in cold water, poorly soluble in hot water, but quite soluble in alcohol and lipids. A quercetin glycoside is formed by attaching a glycosyl group (a sugar such as glucose, rhamnose, or rutinose) as a replacement for one of the OH groups (commonly at position 3). The attached glycosyl group can change the solubility, absorption, and in vivo effects. As a general rule of thumb, the presence of a glycosyl group (quercetin glycoside) results in increased water solubility compared to quercetin aglycone 18.
A quercetin glycoside is unique by the attached glycosyl group. Generally, the term quercetin should be used to describe the aglycone only; however, the name is occasionally used to refer to quercetin-type molecules, including its glycosides in research and the supplement industry.
Quercetin Absorption
Quercetin glycosides might be differently absorbed based on the type of sugar attached 19. Available evidence indicates that quercetin glucosides (like those found predominantly in onion or shallot flesh) are far better absorbed than its rutinosides (the major quercetin glycoside in tea). The glucosides are efficiently hydrolyzed in the small intestine by beta-glucosidases to the aglycone form, much of which is then absorbed 20. Quercetin glucuronic acid and its sulfuric acid derivatives were more easily absorbed than quercetin 20. Thereafter, its absorption is affected by differences in its glycosylation, the food matrix from which it is consumed, and the co-administration of dietary components such as fiber and fat 21. Thus different sugar types and sugar group conjugation sites will result in absorption variation.
Quercetin and derivatives are stable in gastric acid; however, there were no reports whether they can be absorbed in stomach. Studies suggest that quercetin is absorbed in the upper segment of small intestinal 22.
Among quercetin’s derivatives, conjugated forms of its glycosides are better absorbed than quercetin. Purified quercetin glucosides are capable of interacting with the sodium dependent glucose transport receptors in the mucosal epithelium and may therefore be absorbed by the small intestine in vivo 23.
Quercetin foods
Quercetin-type flavonols (primarily as quercetin glycosides), the most abundant of the flavonoid molecules, are widely distributed in plants. They are found in a variety of foods including apples, berries, Brassica vegetables, capers, grapes, onions, shallots, tea, and tomatoes, as well as many seeds, nuts, flowers, barks, and leaves. Quercetin is also found in medicinal botanicals, including Ginkgo biloba, Hypericum perforatum, and Sambucus canadensis 24, 25, 26. In red onions, higher concentrations of quercetin occur in the outermost rings and in the part closest to the root, the latter being the part of the plant with the highest concentration 27. One study found that organically grown tomatoes had 79% more quercetin than chemically grown fruit 28. Quercetin is present in various kinds of honey from different plant sources 29. Food-based sources of quercetin include vegetables, fruits, berries, nuts, beverages and other products of plant origin 30. In the determined food, the highest concentration is 234 mg/100 g of edible portion in capers (raw), the lowest concentration is 2 mg/100 g of edible portion in black or green tea (Camellia sinensis) 31.
Dietary intake of quercetin was different in several countries. The estimated flavonoid intake ranges from 50 to 800 mg/day (quercetin accounts for 75%), mostly depending on the consumption of fruits and vegetables and the intake of tea 32. In the Suihua area of northern China, quercetin intake was reported to be 4.37 mg/day, where the main food sources of flavonol was apples (7.4%), followed by potatoes (3.9%), lettuce (3.8%) and oranges (3.8%) 33, whereas the average quercetin intake was 4.43 mg/day, with apple (3.7%), potato (2.5%), celery (2.2%), eggplant (2.2%), and actinidia (1.6%) being the main food sources of quercetin in Harbin, China 34. The most recent study showed that quercetin intake is about 18 mg/day for Chinese healthy young males. In the USA, flavonol intake is about 13 mg/day for U.S. adults, while quercetin represents three-quarters of this amount. The mean quercetin intake was approximately 14.90 to 16.39 mg per day. Onions, tea, and apples contained high amounts of quercetin 35. In Japan, the average and median quercetin intakes were 16.2 and 15.5 mg/day, respectively; the quercetin intake by men was lower than that by women; and the quercetin intakes showed a low correlation with age in both men and women. The estimated quercetin intake was similar during summer and winter. Quercetin was mainly ingested from onions and green tea, both in summer and in winter. Vegetables, such as asparagus, green pepper, tomatoes, and red leaf lettuce, were good sources of quercetin in summer 36. In Australia, black and green teas were the dominant sources of quercetin. Other sources included onion, broccoli, apple, grape, and beans 37. Analysis of the 24-h recall data indicated an average adult intake of total flavonoids (>18 years) of 454 mg/day. Apple was the most important source of quercetin until age 16–18 years, after which onion became an increasingly important prominent source 37. In Spain, the average daily intake of quercetin is 18.48 mg/day, which is significantly higher than that in the United States (9.75 mg/day), based on sources like tea, citrus fruits and juice, beers and ales, wines, melon, apples, onions, berries and bananas 38.
Table 1. Foods containing quercetin
| Foods containing quercetin | Quercetin (mg/100g of edible portion) |
|---|---|
| capers, raw | 234 |
| capers, canned | 173 |
| lovage | 170 |
| dock like sorrel | 86 |
| radish leaves | 70 |
| carob fiber | 58 |
| dill | 55 (48-110) |
| cilantro | 53 |
| Hungarian wax pepper | 51 |
| fennel leaves | 48.8 |
| onion, red | 32 |
| radicchio | 31.5 |
| watercress | 30 |
| buckwheat | 23 |
| kale | 23 |
| chokeberry | 19 |
| cranberry | 15 |
| lingonberry | 13 |
| plums, black | 12 |
| cow peas | 11 |
| sweet potato | 10 |
| blueberry, cultivated | 8 |
| sea buckthorn berry | 8 |
| rowanberry | 7 |
| crowberry | 5 |
| prickly pear cactus fruits | 5 |
| apples, Red Delicious | 4 |
| broccoli | 3 |
| bilberry | 3 |
| tea, black or green Camellia sinensis | 2 |
| red kidney beans, raw (powdered) | 0.0603 +/- 0.0307 |
Quercetin benefits
Pharmacological reports have demonstrated that quercetin has pronounced pharmaceutical effects, including anti-inflammatory, antioxidant, and antiviral activity 41. In addition, quercetin is included in a novel class of chemotherapeutic drugs for the treatment of various cancers 42 and can also be combined with ultrasonic pretreatment to increase the concentration of quercetin to inhibit the growth of the prostate and skin cancer cell lines 43. However, quercetin has low aqueous solubility, poor absorption, and rapid metabolism (bioavailability approximately 1%–5%) 44, all of which can generate in vivo (animal & human study) results that differ from the powerful in vitro (test tube study) efficacy of quercetin. Most of the studies which have shown the antitumor activity of quercetin were performed with a high concentration of quercetin, rangin from 25 μM to 200 μM. But pharmacokinetic research reveals that the peak concentration of quercetin in blood after food uptake reaches to a peak of around 10 μM 45.
Quercetin has been shown to inhibit the proliferation of a wide range of cancers such as prostate, cervical, lung, breast, and colon. Recent studies have revealed that quercetin inhibits cell proliferation by causing apoptosis and/or cell cycle arrest 46. It has been shown that quercetin treatment causes cell cycle arrests such as G2/M arrest or G1 arrest in different cell types 47. Moreover, quercetin-mediated apoptosis may result from the induction of stress proteins, disruption of microtubules and mitochondria, release of cytochrome c, and activation of caspases 48. Quercetin is a strong antioxidant because it can chelate metals, scavenge oxygen free radicals 49 and inhibit xanthine oxidase and lipid peroxidation in vitro 50.
Quercetin has also been shown to impair an oncoviral replication. Quercetin inhibited HBsAg (hepatitis B surface antigen) and HBeAg (hepatitis B e antigen) secretion in the HBV (Hepatitis virus B)-producing 2.2.15 cells 51. In addition, quercetin appeared to be the most effective inhibitor for HCV (hepatitis C virus) replication among all flavonoids, demonstrating a strong anti-hepatitis C virus activity in hepatitis C virus replicon-containing cells when combined with interferon (IFN)α 52. Quercetin and its analog quercetin showed stronger inhibition on HIV-1 reverse transcriptase, all with IC50 values of 60 μM than HIV-1 protease and α-Glucosidase 53. Quercetin was shown to inhibit other three reverse transcriptases from avian myeloblastosis, Rous-associated virus-2 and Maloney murine leukemia virus when poly(rA)oligo(dT)12-18 or rabbit globin mRNA were used as template 54. Several studies investigated potential use for quercetin as anti-cancer agent 55. A couple of cell culture studies showed that quercetin has an anti-cancer activity due to its antioxidant or anti-inflammatory properties 56. Quercetin also inhibits the growth of cancer cells and helps induction of apoptosis 57. Some studies using animal models have shown that quercetin could protect against colon cancer 58.
Quercetin Clinical Studies
Diet supplementation with combinations of resveratrol, pterostilbene, morin hydrate, quercetin, δ-tocotrienol, riboflavin, and nicotinic acid reduces cardiovascular risk factors in humans when used as nutritional supplements with, or without, other dietary changes in healthy seniors and hypercholesterolemic subjects 59.
In a randomized, double-blinded, placebo-controlled trial, 1002 subjects took 500 or 1000 mg/day quercetin or a placebo for 12 weeks. For the group as a whole, quercetin supplementation had no significant influence on rates of upper respiratory tract infections (URTI) compared to placebo. In a subgroup of subjects age 40 or older who self-rated themselves as physically fit, 1000 mg/day quercetin resulted in a statistically significant reduction in total sick days and symptom severity associated with URTI 60. Female subjects were supplemented with 500 or 1000 mg/day quercetin or placebo for 12 weeks. While quercetin supplementation significantly increased plasma quercetin levels, it had no influence on measure of immune function 61. Quercetin (100 mg/day) did not alter exercise-induced changes in several measures of immune function following three days of intense exercise in trained athletes, but it significantly reduced URTI incidence (1 of 20 subjects in active versus 9 of 20 in placebo group) during the two-week post-exercise period 62. A similar lack of effect on strenuous exercise-induced immune system perturbation was found in subjects who took 1000 mg/day of quercetin for three weeks before, during, and continuing for two weeks after the 160-km Western States Endurance Run. In this study, however, there were no differences in the post-race illness rates between quercetin and placebo groups 63.
There are several studies in humans investigating the correlation of quercetin and its immunomodulatory effects. Quercetin does indeed reduce illness after intensive exercise. Again, under double-blind conditions, Nieman et al. showed that a supplement of 1000 mg of quercetin alone three weeks before, during and two weeks after a three-day period of 3 h of cycling in the winter resulted in a markedly lower incidence of URTI in well-trained subjects in the two weeks after the intensified training, but had no effect on exercise-induced immune dysfunction, inflammation and oxidative stress 64.
The literature is supportive of the anti-pathogenic capacities of quercetin when it is cultured with target cells and a broad spectrum of pathogens including URTI-related rhinoviruses, adenoviruses and coronaviruses. The impact of the co-ingestion of two or more flavonoids increases their bioavailability and the outcomes on immunity. Nieman et al. determined the influence of two weeks of 1000 mg/day quercetin compared with placebo supplementation on exercise performance and skeletal muscle mitochondrial biogenesis in untrained, young adult males. It resulted in significantly reduced post-exercise measures for both inflammation and oxidative stress, with a chronic augmentation of granulocyte oxidative burst activity 65. When taken together, quercetin showed a successful reduction in the illness rates of exercise-stressed athletes as well as a chronic augmentation of their innate immune function.
Most in vitro research suggests that quercetin possesses anti-inflammation and immunological improvement. However, the results from a double-blinded, placebo-controlled, randomized trial indicated that quercetin supplementation at 500 and 1000 mg/day for 12 weeks significantly increased plasma quercetin levels but had no influence on measures of innate immune function or inflammation in community-dwelling adult females 61.
The main action of quercetin on inflammation and immune function in vivo is summarized in the Table 2.
- These results suggest that quercetin studies in human did not totally support these results from cells and animals. The effect, in which quercetin acts as an immune booster in humans, needs to be further verified for future broad application.
Table 2. Summary of the main effects of quercetin on inflammation and immune function in human
| Dosage | Subjects | Effect | Mechanism | Reference |
|---|---|---|---|---|
| Human | ||||
| 50 and 100 mg/person | Elderly Human subject | Anti-inflammatory properties | Inhibition of proteasome (nitric oxide, C-reactive protein, γ-glutamyltransferase) activity | 66 |
| 500 and 1000 mg/day | Human subject | Reduction of upper respiratory tract infection and total sick days; Improvement in 12-min treadmill time trial performance | No effect | 67 |
| 1000 mg/day | Human in treadmill | No effect | 68 | |
| 500 and 1000 mg/day | Human subject | No effect on innate immune function or inflammation, illness rates | No effect | 69 |
| 1000 mg/day | Human cyclist | No effect | 70 | |
| 1000 mg/day | Human runner | No effect | 71 | |
| 1000 mg/day | Human cyclist | No effect | 72 | |
Quercetin side effects
Evaluation: There is inadequate evidence in humans for the carcinogenicity of quercetin. There is limited evidence in experimental animals for the carcinogenicity of quercetin 73. Groups of 50 male and 50 female Fischer 344/DuCrj rats, six weeks of age, were fed quercetin (purity, at least 99.4%) in the diet at concentrations of 0 (control), 1.25 or 5% for 104 weeks and were maintained for a further eight weeks without quercetin. The high dose was the maximum tolerated. At the end of the 112-week study, the survival rates for males were 56, 66 and 68% and those for females were 66, 62 and 72% for the three groups, respectively. No statistically significant increase in incidence of tumors was seen, but males at the high dose showed a significant increase in the incidence of non-neoplastic hyperplastic polyps of the cecum, and one adenoma and two adenocarcinomas of
the cecum were found in males at this dose and two adenomas of the colon occurred in females 74.
Groups of 70 male and 70 female Fischer 344/N rats, seven weeks of age, were given quercetin (purity, > 95%; ellagic acid was the predominant impurity at 1.1–2.6%) in the diet at concentrations of 0, 1000, 10 000 or 40 000 mg/kg (ppm) for 104 weeks. Ten animals per group were killed at 6 and 15 months. The high dose approached the maximum tolerated. Treatment did not affect the survival of either male or female rats, but the decreased body-weight gain of animals at the high dose was attributed to quercetin. Males at the high-dose had an increased incidence of renal tubular tumours (three adenomas and one adenocarcinoma) with none in control males, but the increase did not achieve statistical significance. After step-sectioning, a total of nine renal tubular tumours were found in these animals, and the increase was statistically significant. The severity of spontaneous progressive nephropathy was exacerbated in male rats by exposure to quercetin 75.
Overall evaluation: Quercetin is not classifiable as to its carcinogenicity to humans (Group 3) 73.
References- Endothelial function and cardiovascular disease: effects of quercetin and wine polyphenols. Free Radic Res. 2006 Oct;40(10):1054-65. https://www.ncbi.nlm.nih.gov/pubmed/17015250
- Committee of Pharmacognosy Publication. Pharmacognosy. Pharmacognosy. Seoul Korea: Dong Myoung; 2013. pp. 102–105.
- Intake of potentially anticarcinogenic flavonoids and their determinants in adults in The Netherlands. Hertog MG, Hollman PC, Katan MB, Kromhout D. Nutr Cancer. 1993; 20(1):21-9. https://www.ncbi.nlm.nih.gov/pubmed/8415127/
- Effects of the dietary flavonoid quercetin upon performance and health. Davis JM, Murphy EA, Carmichael MD. Curr Sports Med Rep. 2009 Jul-Aug; 8(4):206-13. https://www.ncbi.nlm.nih.gov/pubmed/19584608/
- Aguirre L., Arias N., Macarulla M.T., Gracia A., Portillo M.P. Beneficial effects of quercetin on obesity and diabetes. Open Nutraceuticals J. 2011;4:189–198.
- National Toxicology Program (1991) NTP Chemical Repository Data Sheet: Quercetin , Research Triangle Park, NC.
- Middleton E Jr, Kandaswami C, Theoharides TC. The effects of plant flavonoids on mammalian cells: implications for inflammation, heart disease and cancer. Pharmacol Rev 2000;52:673–751.
- Beecher GR. Overview of dietary flavonoids: nomenclature, occurrence and intake. J Nutr 2003;133:3248S–54S.
- Del Rio D, Rodriguez-Mateos A, Spencer JP, Tognolini M, Borges G, Crozier A. Dietary (poly)phenolics in human health: structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxid Redox Signal 2013;18:1818–92.
- Russo M, Spagnuolo C, Tedesco I, Bilotto S, Russo GL. The flavonoid quercetin in disease prevention and therapy: facts and fancies. Biochem Pharmacol 2012;83:6–15.
- Roskosko R Jr. ERK1/2 MAP kinases: structure, function and regulation. Pharmacol Res 2012;66:105–43.
- Gutiérrez-Venegas G, Bando-Campos CG. The flavonoids luteolin and quercetagetin inhibit lipoteichoic acid actions on H9c2 cardiomyocytes. Int Immunopharmacol 2010;10:1003–9.
- Walker EH, Pacold ME, Perisic O, Stephens L, Hawkins PT, Wymann MP et al. Structural determinants of phosphoinositide 3-kinase inhibition by wortmannin, LY294002, quercetin, myricetin, and staurosporine. Mol Cell 2000;6:909–19.
- Hausenloy DJ, Yellon DM. Preconditioning and postconditioning: united at reperfusion. Pharmacol Ther 2007;114:208–21.
- Shao Z-H, Wojcik KR, Dossumbekova A, Hsu C, Mehendale SR, Li C-Q et al. Grape seed proanthocyanidins protect cardiomyocytes from ischemia and reperfusion injury via Akt-NOS signaling. J Cell Biochem 2009;107:697–705.
- Angeloni C, Spencer JPE, Leoncini E, Biagi PL, Hrelia S. Role of quercetin and its in vivo metabolites in protecting H9c2 cells against oxidative stress. Biochimie 2007;89:73–82.
- Sun B, Sun GB, Xiao J, Chen RC, Wang X, Wu Y et al. Isorhamnetin inhibits H2O2-induced activation of the intrinsic apoptotic pathway in H9c2 cardiomyocytes through scavenging reactive oxygen species and ERK inactivation. J Cell Biochem 2012;113:473–85.
- Dietary flavonoids: bioavailability, metabolic effects, and safety. Ross JA, Kasum CM. Annu Rev Nutr. 2002; 22():19-34. https://www.ncbi.nlm.nih.gov/pubmed/12055336/
- Scholz S., Williamson G. Interactions affecting the bioavailability of dietary polyphenols in vivo. Int. J. Vitam. Nutr. Res. 2007;77:224–235. doi: 10.1024/0300-9831.77.3.224. https://www.ncbi.nlm.nih.gov/pubmed/18214024
- Ader P., Wessmann A., Wolffram S. Bioavailability and metabolism of the flavonol quercetin in the pig. Free Radic. Biol. Med. 2000;28:1056–1067. doi: 10.1016/S0891-5849(00)00195-7.
- Guo Y., Mah E., Davis C.G., Jalili T., Ferruzzi M.G., Chun O.K., Bruno R.S. Dietary fat increases quercetin bioavailability in overweight adults. Mol. Nutr. Food Res. 2013;57:896–905. doi: 10.1002/mnfr.201200619.
- Crespy V., Morand C., Manach C. Part of quercetin absorbed in the small intestine is conjugated and further secreted in the intestinal l: Umen. Am. J. Physiol. 1999;277:G120–G126.
- Scholz S., Williamson G. Interactions affecting the bioavailability of dietary polyphenols in vivo. Int. J. Vitam. Nutr. Res. 2007;77:224–235. doi: 10.1024/0300-9831.77.3.224.
- Wiczkowski W., Romaszko J., Bucinski A., Szawara-Nowak D., Honke J., Zielinski H., Piskula M.K. Quercetin from shallots (Allium cepa L. var. aggregatum) is more bioavailable than its glucosides. J. Nutr. 2008;138:885–888. http://jn.nutrition.org/content/138/5/885.long
- Häkkinen S.H., Kärenlampi S.O., Heinonen I.M., Mykkänen H.M., Törrönen A.R. Content of the flavonols quercetin, myricetin, and kaempferol in 25 edible berries. J. Agric. Food Chem. 1999;47:2274–2279. doi: 10.1021/jf9811065.
- Williamson G., Manach C. Bioavailability and bioefficacy of polyphenols in humans. II. Review of 93 intervention studies. Am. J. Clin. Nutr. 2005;81(Suppl. S1):243S–255S.
- Smith C., Lombard K.A., Peffley E.B., Liu W. Genetic analysis of quercetin in onion (Allium cepa L.) Lady Raider. Tex. J. Agric. Natl. Resour. 2003;16:24–28.
- Mitchell A.E., Hong Y.J., Koh E., Barrett D.M., Bryant D.E., Denison R.F., Kaffka S. Ten-year comparison of the influence of organic and conventional crop management practices on the content of flavonoids in tomatoes. J. Agric. Food Chem. 2007;55:6154–6159. doi: 10.1021/jf070344+.
- Petrus K., Schwartz H., Sontag G. Analysis of flavonoids in honey by HPLC coupled with coulometric electrode array detection and electrospray ionization mass spectrometry. Anal. Bioanal. Chem. 2011;400:2555–2563. doi: 10.1007/s00216-010-4614-7.
- Tutelian V.A., Lashneva N.V. Biologically active substances of plant origin. Flavonols and flavones: Prevalence, dietary sources and consumption. Vopr. Pitan. 2013;82:4–22.
- Bhagwat S., Haytowits D.B., Holden J.M. USDA Database for the Flavonoid Content of Selected Foods, Release 3. U.S. Department of Agriculture; Beltsville, MD, USA: 2011.
- Chun O.K., Chung S.J., Song W.O. Estimated dietary flavonoid intake and major food sources of U.S. adults. J. Nutr. 2007;137:1244–1252.
- Sun C., Wang H., Wang D., Chen Y., Zhao Y., Xia W. Using an FFQ to assess intakes of dietary flavonols and flavones among female adolescents in the Suihua area of northern China. Public Health Nutr. 2015;18:632–639. doi: 10.1017/S1368980014000780.
- Zhang Y., Li Y., Cao C., Cao J., Chen W., Zhang Y., Wang C., Wang J., Zhang X., Zhao X. Dietary flavonol and flavone intakes and their major food sources in Chinese adults. Nutr. Cancer. 2010;62:1120–1127. doi: 10.1080/01635581.2010.513800.
- Sampson L., Rimm E., Hollman P.C., de Vries J.H., Katan M.B. Flavonol and flavone intakes in US health professionals. J. Am. Diet. Assoc. 2002;102:1414–1420. doi: 10.1016/S0002-8223(02)90314-7.
- Nishimuro H., Ohnishi H., Sato M., Ohnishi-Kameyama M., Matsunaga I., Naito S., Ippoushi K., Oike H., Nagata T., Akasaka H., et al. Estimated daily intake and seasonal food sources of quercetin in Japan. Nutrients. 2015;7:2345–2358. doi: 10.3390/nu7042345.
- Somerset S.M., Johannot L. Dietary flavonoid sources in Australian adults. Nutr. Cancer. 2008;60:442–449. doi: 10.1080/01635580802143836.
- Zamora-Ros R., Andres-Lacueva C., Lamuela-Raventos R.M., Berenguer T., Jakszyn P., Barricarte A., Ardanaz E., Amiano P., Dorronsoro M., Larrañaga N., et al. Estimation of dietary sources and flavonoid intake in a Spanish adult population (EPIC-Spain) J. Am. Diet. Assoc. 2010;110:390–398. doi: 10.1016/j.jada.2009.11.024.
- USDA Database for the Flavonoid Content of Selected Foods Release 3, September 2011. https://www.ars.usda.gov/ARSUserFiles/80400525/Data/Flav/Flav_R03.pdf
- Justesen U, Knuthsen P (May 2001). “Composition of flavonoids in fresh herbs and calculation of flavonoid intake by use of herbs in traditional Danish dishes”. Food Chemistry. 73 (2): 245–50. doi:10.1016/S0308-8146(01)00114-5.
- Kaushik D, O’Fallon K, Clarkson PM, Dunne CP, Conca KR, Michniak-Kohn B. Comparison of quercetin pharmacokinetics following oral supplementation in humans. J Food Sci. 2012;77(11):H231–H238.
- Lee SM, Moon J, Chung JH, Cha YJ, Shin MJ. Effect of quercetin-rich onion peel extracts on arterial thrombosis in rats. Food Chem Toxicol. 2013;57:99–105.
- Paliwal S, Sundaram J, Mitragotri S. Induction of cancer-specific cytotoxicity towards human prostate and skin cells using quercetin and ultrasound. Br J Cancer. 2005;92(3):499–502.
- Reinboth M, Wolffram S, Abraham G, Ungemach FR, Cermak R. Oral bioavailability of quercetin from different quercetin glycosides in dogs. Br J Nutr. 2010;104(2):198–203.
- Relative bioavailability of the antioxidant flavonoid quercetin from various foods in man. Hollman PC, van Trijp JM, Buysman MN, van der Gaag MS, Mengelers MJ, de Vries JH, Katan MB. FEBS Lett. 1997 Nov 24; 418(1-2):152-6.
- Quercetin arrests G2/M phase and induces caspase-dependent cell death in U937 cells. Lee TJ, Kim OH, Kim YH, Lim JH, Kim S, Park JW, Kwon TK. Cancer Lett. 2006 Aug 28; 240(2):234-42. https://www.ncbi.nlm.nih.gov/pubmed/16274926/
- Quercetin-induced growth inhibition and cell death in nasopharyngeal carcinoma cells are associated with increase in Bad and hypophosphorylated retinoblastoma expressions. Ong CS, Tran E, Nguyen TT, Ong CK, Lee SK, Lee JJ, Ng CP, Leong C, Huynh H. Oncol Rep. 2004 Mar; 11(3):727-33. https://www.ncbi.nlm.nih.gov/pubmed/14767529/
- Quercetin inhibits Shc- and phosphatidylinositol 3-kinase-mediated c-Jun N-terminal kinase activation by angiotensin II in cultured rat aortic smooth muscle cells. Yoshizumi M, Tsuchiya K, Kirima K, Kyaw M, Suzaki Y, Tamaki T. Mol Pharmacol. 2001 Oct; 60(4):656-65.
- Free radical scavenging and antioxidant activity of plant flavonoids. Kandaswami C, Middleton E Jr. Adv Exp Med Biol. 1994; 366():351-76. https://www.ncbi.nlm.nih.gov/pubmed/7771265/
- Inhibition of the metmyoglobin-induced peroxidation of linoleic acid by dietary antioxidants: Action in the aqueous vs. lipid phase. Vulcain E, Goupy P, Caris-Veyrat C, Dangles O. Free Radic Res. 2005 May; 39(5):547-63.
- In vivo and in vitro antiviral activity of hyperoside extracted from Abelmoschus manihot (L) medik. Wu LL, Yang XB, Huang ZM, Liu HZ, Wu GX. Acta Pharmacol Sin. 2007 Mar; 28(3):404-9.
- Modulation of PI3K-LXRα-dependent lipogenesis mediated by oxidative/nitrosative stress contributes to inhibition of HCV replication by quercetin. Pisonero-Vaquero S, García-Mediavilla MV, Jorquera F, Majano PL, Benet M, Jover R, González-Gallego J, Sánchez-Campos S. Lab Invest. 2014 Mar; 94(3):262-74.
- Effects of triterpenoids and flavonoids isolated from Alnus firma on HIV-1 viral enzymes. Yu YB, Miyashiro H, Nakamura N, Hattori M, Park JC. Arch Pharm Res. 2007 Jul; 30(7):820-6.
- Inhibition of reverse transcriptases by flavonoids. Spedding G, Ratty A, Middleton E Jr. Antiviral Res. 1989 Sep; 12(2):99-110.
- Xavier CP, Lima CF, Rohde M, Pereira-Wilson C. Quercetin enhances 5-fluorouracil-induced apoptosis in MSI colorectal cancer cells through p53 modulation. Cancer chemotherapy and pharmacology. 2011;68:1449–1457.
- Dajas F. Life or death: neuroprotective and anticancer effects of quercetin. Journal of ethnopharmacology. 2012;143:383–396.
- Murphy EA, Davis JM, McClellan JL, Carmichael MD. Quercetin’s effects on intestinal polyp multiplicity and macrophage number in the Apc(Min/+) mouse. Nutrition and cancer. 2011;63:421–426.
- Sekine-Osajima Y, Sakamoto N, Nakagawa M, Itsui Y, Tasaka M, Nishimura-Sakurai Y, Chen CH, Suda G, Mishima K, Onuki Y, Yamamoto M, Maekawa S, Enomoto N, Kanai T, Tsuchiya K, Watanabe M. Two flavonoids extracts from Glycyrrhizae radix inhibit in vitro hepatitis C virus replication. Hepatology research : the official journal of the Japan Society of Hepatology. 2009;39:60–69.
- Qureshi A.A., Khan D.A., Mahjabeen W., Papasian C.J., Qureshi N. Suppression of nitric oxide production and cardiovascular risk factors in healthy seniors and hypercholesterolemic subjects by a combination of polyphenols and vitamins. J. Clin. Exp. Cardiol. 2012;S5:167. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3486425/
- Heinz S.A., Henson D.A., Austin M.D., Jin F., Nieman D.C. Quercetin supplementation and upper respiratory tract infection: A randomized community clinical trial. Pharmacol. Res. 2010;62:237–242. doi: 10.1016/j.phrs.2010.05.001. https://www.ncbi.nlm.nih.gov/pubmed/20478383
- Heinz S.A., Henson D.A., Nieman D.C., Austin M.D., Jin F. A 12-week supplementation with quercetin does not affect natural killer cell activity, granulocyte oxidative burst activity or granulocyte phagocytosis in female human subjects. Br. J. Nutr. 2010;104:849–857. doi: 10.1017/S000711451000156X. https://www.ncbi.nlm.nih.gov/pubmed/20500927
- Nieman D.C., Henson D.A., Gross S.J., Jenkins D.P., Davis J.M., Murphy E.A., Carmichael M.D., Dumke C.L., Utter A.C., McAnulty S.R., et al. Quercetin reduces illness but not immune perturbations after intensive exercise. Med. Sci. Sports Exerc. 2007;39:1561–1569. doi: 10.1249/mss.0b013e318076b566. https://www.ncbi.nlm.nih.gov/pubmed/17805089
- Henson D., Nieman D., Davis J.M., Dumke C., Gross S., Murphy A., Carmichael M., Jenkins D.P., Quindry J., McAnulty S., et al. Post-160-km race illness rates and decreases in granulocyte respiratory burst and salivary IgA output are not countered by quercetin ingestion. Int. J. Sports Med. 2008;29:856–863. doi: 10.1055/s-2007-989424. https://www.ncbi.nlm.nih.gov/pubmed/18213545
- Nieman D.C., Henson D.A., Maxwell K.R., Williams A.S., McAnulty S.R., Jin F., Shanely R.A., Lines T.C. Effects of quercetin and EGCG on mitochondrial biogenesis and immunity. Med. Sci. Sports Exerc. 2009;41:1467–1475. doi: 10.1249/MSS.0b013e318199491f. https://www.ncbi.nlm.nih.gov/pubmed/19516153
- Nieman D.C., Williams A.S., Shanely R.A., Jin F., McAnulty S.R., Triplett N.T., Austin M.D., Henson D.A. Quercetin’s influence on exercise performance and muscle mitochondrial biogenesis. Med. Sci. Sports Exerc. 2010;42:338–345. doi: 10.1249/MSS.0b013e3181b18fa3. https://www.ncbi.nlm.nih.gov/pubmed/19927026
- Qureshi A.A., Khan D.A., Mahjabeen W., Papasian C.J., Qureshi N. Suppression of nitric oxide production and cardiovascular risk factors in healthy seniors and hypercholesterolemic subjects by a combination of polyphenols and vitamins. J. Clin. Exp. Cardiol. 2012;S5:167.
- Heinz S.A., Henson D.A., Austin M.D., Jin F., Nieman D.C. Quercetin supplementation and upper respiratory tract infection: A randomized community clinical trial. Pharmacol. Res. 2010;62:237–242. doi: 10.1016/j.phrs.2010.05.001.
- Nieman D.C., Henson D.A., Maxwell K.R., Williams A.S., McAnulty S.R., Jin F., Shanely R.A., Lines T.C. Effects of quercetin and EGCG on mitochondrial biogenesis and immunity. Med. Sci. Sports Exerc. 2009;41:1467–1475. doi: 10.1249/MSS.0b013e318199491f.
- Heinz S.A., Henson D.A., Nieman D.C., Austin M.D., Jin F. A 12-week supplementation with quercetin does not affect natural killer cell activity, granulocyte oxidative burst activity or granulocyte phagocytosis in female human subjects. Br. J. Nutr. 2010;104:849–857. doi: 10.1017/S000711451000156X.
- Nieman D.C., Henson D.A., Gross S.J., Jenkins D.P., Davis J.M., Murphy E.A., Carmichael M.D., Dumke C.L., Utter A.C., McAnulty S.R., et al. Quercetin reduces illness but not immune perturbations after intensive exercise. Med. Sci. Sports Exerc. 2007;39:1561–1569. doi: 10.1249/mss.0b013e318076b566.
- Henson D., Nieman D., Davis J.M., Dumke C., Gross S., Murphy A., Carmichael M., Jenkins D.P., Quindry J., McAnulty S., et al. Post-160-km race illness rates and decreases in granulocyte respiratory burst and salivary IgA output are not countered by quercetin ingestion. Int. J. Sports Med. 2008;29:856–863. doi: 10.1055/s-2007-989424.
- Nieman D.C., Williams A.S., Shanely R.A., Jin F., McAnulty S.R., Triplett N.T., Austin M.D., Henson D.A. Quercetin’s influence on exercise performance and muscle mitochondrial biogenesis. Med. Sci. Sports Exerc. 2010;42:338–345. doi: 10.1249/MSS.0b013e3181b18fa3.
- https://monographs.iarc.fr/ENG/Monographs/vol73/mono73-23.pdf
- Ito, N., Hagiwara, A., Tamano, S., Kagawa, M., Shibata, M.-A., Kurata, Y. & Fukushima, S.(1989) Lack of carcinogenicity of quercetin in F344/DuCrj rats. Jpn. J. Cancer Res .,80,317–325.
- National Toxicology Program (1992). Toxicology and Carcinogenesis Studies of Quercetin (CASNo. 117-39-5) in F344/N Rats (Feed Studies) (Tech. Rep. Ser. No. 409; NIH Publ. No. 92-3140), Research Triangle Park, NC.



{kind=link}